Microbiologia Ambiental
Prezado(a) aluno(a), seja bem-vindo(a) à Unidade I da disciplina de Microbiologia. A Microbiologia é a ciência que tem por objetivo estudar os micro-organismos, nos quais se incluem bactérias, fungos, vírus e protozoários. Nesta unidade, dedicaremo-nos a conhecer como os micro-organismos foram descobertos e quais os principais grupos desses organismos. Também estudaremos a morfologia, a classificação e a reprodução desses organismos. No decorrer desta unidade, veremos que esses pequenos organismos, que estão associados com doenças e prejuízos, também podem ser utilizados de forma benéfica e possuem inúmeras aplicações, como a produção de alimentos, de fármacos e o controle de pragas. Desejamos que você, caro(a) aluno(a), aproveite ao máximo seus estudos!
No início desta unidade, estudaremos o histórico de descobrimento dos micro-organismos, mencionando os principais acontecimentos, os cientistas e os experimentos que levaram ao desenvolvimento da microbiologia como uma ciência.
Dando início ao nosso estudo, falaremos sobre como aconteceu a descoberta dos micro-organismos. Esta está associada à invenção do microscópio. Em 1665, o inglês Robert Hooke, ao observar uma fatia de cortiça em um microscópio bem rudimentar, descreveu estruturas de frutificação de bolores e declarou que as menores unidades estruturais da vida eram “pequenas caixas” ou “células”. Assim, com a descoberta de Hooke, teve início a teoria celular – em que todas as coisas vivas são formadas por células (MADIGAN et al., 2016; TORTORA; FUNKE; CASE, 2017).
No entanto, o microscópio de Hooke não tinha resolução suficiente para visualizar os micro-organismos com detalhes. Então, em 1684, o comerciante holandês, cientista amador e construtor de microscópios Anton Van Leeuwenhoek foi a primeira pessoa a observar com mais detalhes os micro-organismos. Os microscópios de Leeuwenhoek eram extremamente simples, no entanto, por eles, Leeuwenhoek visualizou e ilustrou micro-organismos (que ele chamou de animálculos) que encontrou em diferentes amostras, como de água de chuva e fezes. Os desenhos de Leeuwenhoek podem ser observados na Figura 1.1. Suas ilustrações foram confirmadas por outros pesquisadores e identificadas como representações de bactérias e protozoários (MADIGAN et al., 2016; TORTORA; FUNKE; CASE, 2017).

Desse modo, o aperfeiçoamento dos microscópios contribuiu com o desenvolvimento da microbiologia, pois permitiu observar, de forma mais precisa, as células bacterianas, o que possibilitou o estudo dos micro-organismos (MADIGAN et al., 2016).
Embora os cientistas Robert Koch e Louis Pasteur sejam lembrados frequentemente quando falamos de microbiologia, Ferdinand Cohn (1828-1898) é o cientista considerado pai da bacteriologia; ele transformou a microbiologia em uma ciência de fato. Cohn era contemporâneo de Pasteur e Koch e havia se especializado em botânica, no entanto, no ano de 1850, interessou-se por microscopia e passou a estudar os processos de crescimento e divisão de células vegetais. Com o tempo, os estudos de Cohn passaram a abranger, além das plantas, algas e bactérias (MADIGAN et al., 2016).
Cohn descobriu o gênero Bacillus, seus estudos o levaram a descrever todo o ciclo desse gênero (célula vegetativa → endósporo → célula vegetativa) e o processo de formação de endósporos, que, hoje, sabemos que são estruturas termorresistentes. Além disso, descobriu que as células vegetativas podiam morrer em água em ebulição, no entanto os endósporos permaneciam viáveis. Assim, essas descobertas ajudaram a entender como os métodos de esterilização (utilizados no passado) que utilizavam água em ebulição nem sempre eram eficientes (MADIGAN et al., 2016).
Os estudos de Cohn colaboraram com o desenvolvimento da bacteriologia como a base de um esquema de classificação de bactérias. Além disso, é atribuído a Cohn participação no desenvolvimento de técnicas simples para evitar a contaminação em meios de cultura, como a utilização de buchas de algodão para fechar tubos e frascos (MADIGAN et al., 2016).
Com a descoberta desses minúsculos organismos por Leeuwenhoek, o mundo científico voltou a sua atenção para a origem deles. Duas questões biológicas intrigaram os cientistas e os filósofos da época, uma dessas questões se referia à geração espontânea, e a outra envolvia a natureza das doenças infecciosas (MADIGAN et al., 2016). Passaremos, a partir agora, a abordar a geração espontânea.
Até metade do século XIX, diversos cientistas e pensadores acreditavam que alguns seres vivos poderiam surgir, espontaneamente, de materiais inanimados. Esse processo hipotético era chamado de geração espontânea. Acreditavam, por exemplo, que larvas surgiam a partir de cadáveres em decomposição e que as moscas tinham origem a partir de estrume (TORTORA; FUNKE; CASE, 2017).
Um dos primeiros passos para contestar essa teoria foi o trabalho realizado por Francesco Redi em 1668. Por meio de seu experimento, Redi tentou demonstrar que as larvas não surgiam espontaneamente. Seu experimento consistia em deixar duas jarras com carne em decomposição. Uma das jarras ficava aberta, o que permitia o livre acesso das moscas ao interior da jarra e à carne, para depositar seus ovos e, posteriormente, o desenvolvimento das larvas. A outra jarra tinha sua abertura selada, assim, as moscas não tinham acesso ao seu interior e, consequentemente nenhuma larva foi observada. No entanto, houve quem não se convenceu com o experimento, argumentando que, para ocorrer a geração espontânea, era necessário ar fresco. Assim, Redi refez o experimento e, ao invés de selar uma das jarras, apenas a cobriu com uma fina rede. Da mesma forma, na jarra que foi coberta com a rede, não pode ser observado o aparecimento de larva. Os resultados dos experimentos de Redi abalaram o conceito de geração espontânea. Contudo alguns cientistas alegaram que formas de vida muito pequenas, como os “animálculos” observados por Leeuwenhoek, eram simples o suficiente para surgirem a partir de matéria inanimada (TORTORA; FUNKE; CASE, 2017).
Em 1858, Rudolf Virchow formulou o conceito de biogênese, contestando a teoria da geração espontânea. Segundo esse conceito, uma célula viva só poderia ter origem a partir de outra célula viva preexistente. Virchow, porém, não conseguiu nenhuma evidência científica para comprovar sua hipótese; assim, a teoria da geração espontânea perdurou até 1861, quando a questão foi finalmente elucidada por Louis Pasteur (TORTORA; FUNKE; CASE, 2017).
O cientista francês Louis Pasteur realizou os trabalhos mais precisos e convincentes, que demonstraram a impossibilidade da geração espontânea. A princípio, Pasteur encheu vários frascos com caldo de carne e os levou à fervura. Ele deixou que alguns desses frascos esfriassem abertos; posteriormente, esses frascos apresentaram crescimento de micróbios. Os outros frascos foram selados após a fervura e não apresentaram crescimento microbiano. A partir desse experimento, Pasteur deduziu que os micróbios do ar eram responsáveis pela contaminação. No entanto os apoiadores da teoria da geração espontânea criticaram o experimento, argumentando que o aquecimento afetava de alguma maneira o ar aprisionado no frasco selado.
Pasteur rebateu as críticas com um segundo experimento, que está ilustrado na Figura 1.2. Ele colocou caldo em frascos com a extremidade longa e afilada e dobrou essa extremidade formando curvas com o formato de um S. O caldo foi fervido e resfriado. Nesse tipo de frasco, as soluções podiam ser fervidas e ele permitia que o ar entrasse livremente após a fervura. Devido ao seu formato, as curvas podiam reter os micróbios que poderiam contaminar a solução. Assim, o conteúdo esterilizado nesses frascos não apresentava contaminação por micróbios e não sofria decomposição (MADIGAN et al., 2016; TORTORA; FUNKE; CASE, 2017).
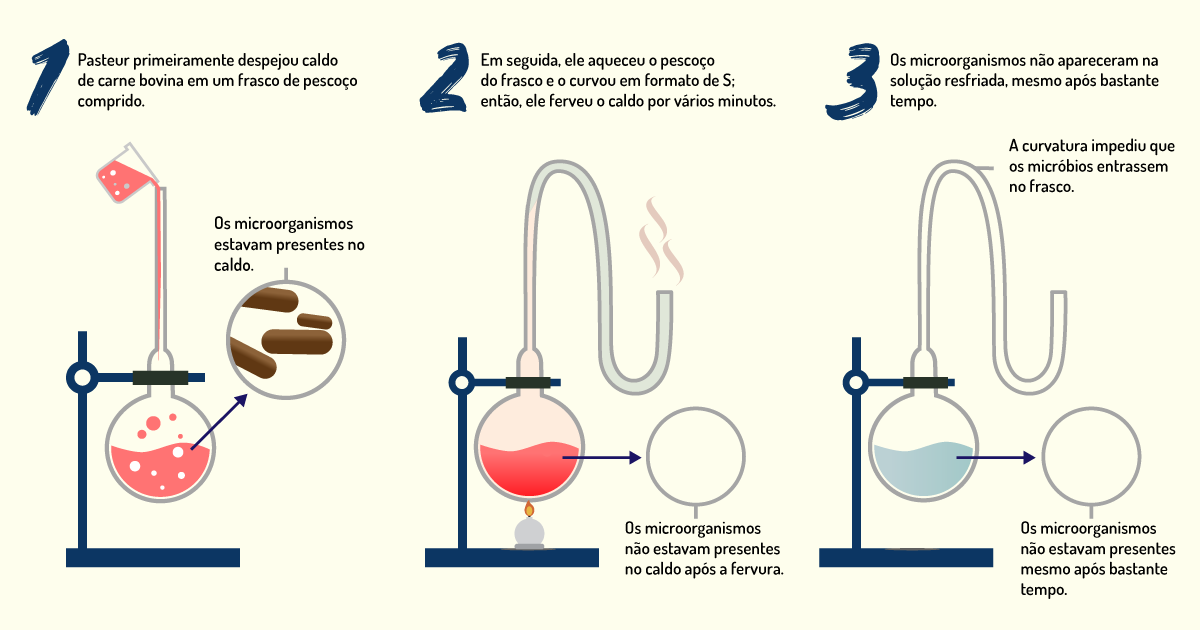
Por meio desses experimentos, Pasteur demonstrou que os micro-organismos podem estar presentes em matéria inanimada, como ar, líquidos e sólidos, e que podem ser destruídos com o calor. Os processos utilizados por Pasteur e outros pesquisadores foram refinados e formam a base das técnicas de assepsia, que eliminam e previnem a contaminação por micro-organismos indesejáveis. Com o fim da teoria da geração espontânea, os procedimentos de esterilização foram refinados e contribuíram com o desenvolvimento da microbiologia como ciência. Além disso, muitos princípios de Pasteur são aplicados à ciência dos alimentos, como no envase e na preservação de diversos tipos de alimentos. Pasteur também demonstrou que a produção de ácido lático e etanol em diferentes tipos de fermentações era resultado da ação dos micro-organismos. Cabe, ainda, destacar que Pasteur desenvolveu vacinas contra as doenças antraz, raiva e cólera aviária. As descobertas de Pasteur na área da medicina e da veterinária também ajudaram a solidificar a teoria do germe e da doença, ou seja, descobriu que as doenças estavam de alguma forma associadas aos germes (MADIGAN et al., 2016; TORTORA; FUNKE; CASE., 2017).
A primeira evidência de que bactérias podem causar doenças foi de Robert Koch em 1876. A princípio, Koch estudou o antraz, uma doença que estava causando a morte dos rebanhos de gado e ovelhas da Europa. Koch verificou que bactérias em forma de bacilo, hoje chamadas de Bacillus anthracis, sempre estavam presentes no sangue do animal que morreu de antraz. A partir dessa observação, Koch percebeu a oportunidade para estudar a doença e utilizou camundongos para realizar seu experimento. Ele observou que, quando uma pequena quantidade de sangue de um camundongo doente era injetada em um camundongo sadio, esse segundo animal apresentava os sintomas da doença. Ao coletar sangue desse segundo camundongo e injetá-lo em um outro camundongo sadio, este também adoecia ou morria. Koch também descobriu que podia cultivar as bactérias em meio de cultura, fora do corpo do animal. Assim, ele injetou as amostras da cultura em animais saudáveis e, mais uma vez, eles apresentaram os sintomas característicos da doença. Koch realizou o isolamento das bactérias presentes no sangue do animal doente e comparou com as bactérias originalmente isoladas e, observou que as duas amostras tinham a mesma bactéria (MADIGAN et al., 2016).
Dessa forma, Koch formulou o Postulado de Koch, que é um conjunto de critérios utilizado para associar um micro-organismo específico a uma doença específica. Assim, esses critérios têm sido utilizados nos últimos 100 anos, para provar que determinado organismo é o agente causador de uma determinada doença. Além disso, as descobertas de Koch enfatizaram a importância da cultura laboratorial do agente infeccioso, o que levou Koch a descobrir os agentes infecciosos de várias doenças. Com isso, o tratamento e a cura dessas doenças foram desenvolvidos, trazendo grande avanço para a área da medicina e bem-estar para o ser humano (MADIGAN et al., 2016).
Caro(a) aluno(a), examinamos, até aqui, os principais fatos históricos que marcaram a história e o desenvolvimento da microbiologia. Passaremos, agora, a estudar a classificação dos micro-organismos, para isso, precisamos entender como todos os organismos, de maneira geral, estão classificados. A taxonomia é um campo científico responsável por classificar os organismos em categorias, ou táxons, de forma a mostrar os graus de semelhança entre eles. Essas semelhanças, por sua vez, estão relacionadas com o parentesco, pois sabemos que os organismos são evolutivamente relacionados. Assim, veremos que a forma como são classificados os organismos passou por muitas mudanças ao longo tempo.
Antes da descoberta dos micróbios, em 1735, os organismos eram distribuídos apenas em reino animal e vegetal, de acordo com o sistema proposto por Carolus Linnaeus. Embora o trabalho de Linnaeus tivesse dado enormes contribuições científicas, seu sistema e os outros sistemas de classificação iniciais eram falhos, pois eram baseados em informações imprecisas (PELCZAR; CHAN; KRIEG, 2011). Com o passar dos anos, vários sistemas foram propostos; como exemplo, em 1800, o sistema de classificação proposto por Carl von Nägeli, agrupava bactérias e fungos no reino vegetal. Posteriormente, em 1959, os fungos foram classificados em seu próprio reino. Em 1937, foi introduzido o conceito de procarioto, que foi utilizado para diferenciar as células anucleadas de células de animais e plantas que são nucleadas (TORTORA; FUNKE; CASE, 2017).
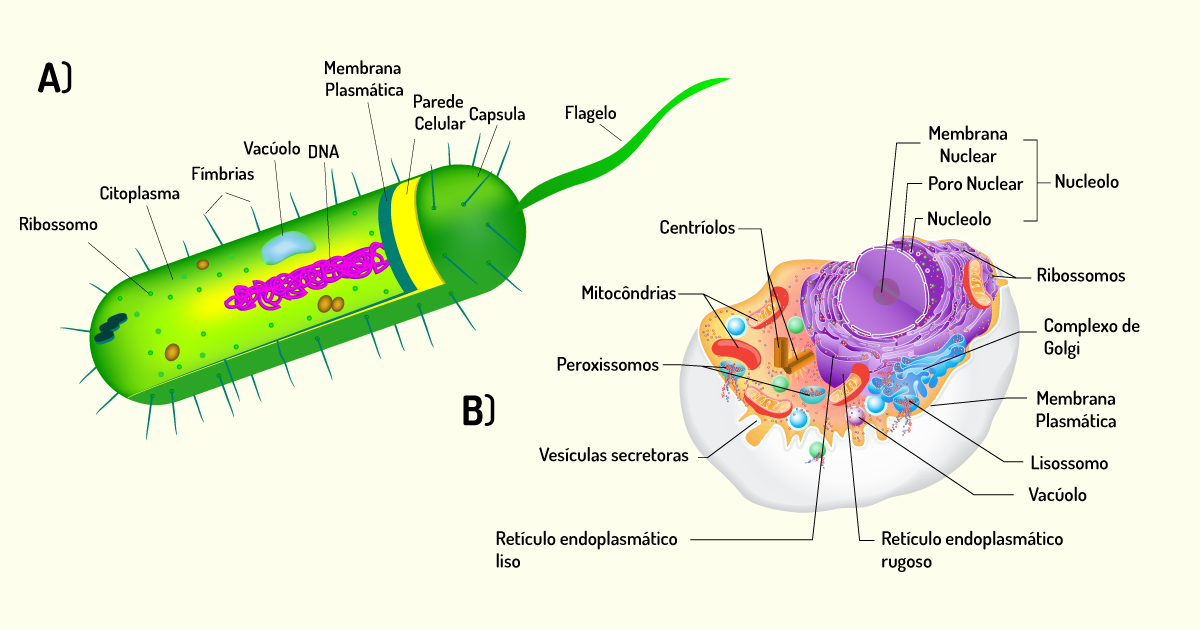
Aqui, caro(a) aluno(a), é importante relembrar as principais diferenças entre procarioto e eucarioto. Organismos chamados de procariotos são unicelulares, ou seja, possuem uma única célula. Além disso, as células desses organismos não possuem um envoltório nuclear, dessa forma, o material genético da célula (DNA) fica em contato direto com o citoplasma, por isso, são chamadas anucleadas (não possuem núcleo). Outra característica é que o DNA dessas células, geralmente, se apresenta como molécula circular. Atualmente, dentre os procariotos, incluem-se as bactérias e as arqueas (bactérias que vivem em ambientes extremos, como lagos com altas concentrações de sal e piscinas térmicas).
Já os organismos eucariotos são mais complexos e, dentre os micro-organismos eucarióticos, estão as algas, os fungos e os protozoários. Todos os organismos pluricelulares, como plantas e animais, são formados por células eucarióticas. Essas células apresentam o DNA envolto por uma membrana e, também, possuem organelas, que são regiões delimitadas por membranas que formam compartimentos e possuem uma função especializada (ZAHA; FERREIRA; PASSAGLIA, 2012; TORTORA; FUNKE; CASE, 2017). As principais diferenças entre as células de procariotos e eucariotos está ilustrada na Figura 1.3.
Prezado(a) aluno(a), relembrados os conceitos das células de procarioto e eucarioto, vamos retomar o sistema de classificação. Vimos que vários sistemas foram propostos ao longo do tempo. Com os avanços na biologia molecular, foi descoberto, que existem dois tipos de células procarióticas e um tipo de célula eucariótica. Essa descoberta, além da análise de sequências de nucleotídeos contidas no RNA ribossomal de diferentes tipos de células, demonstrou que existem três grupos de células: os eucariotos e dois tipos de procariotos (as bactérias e as arqueas) (TORTORA; FUNKE; CASE, 2017).
Então, em 1978, Carl Woese propôs um sistema de classificação baseado na organização celular dos organismos, propondo elevar a um nível acima dos reinos os três tipos de células, nível esse chamado de domínio. A partir de então, os organismos passaram a ser distribuídos em três domínios (conforme Figura 1.4):
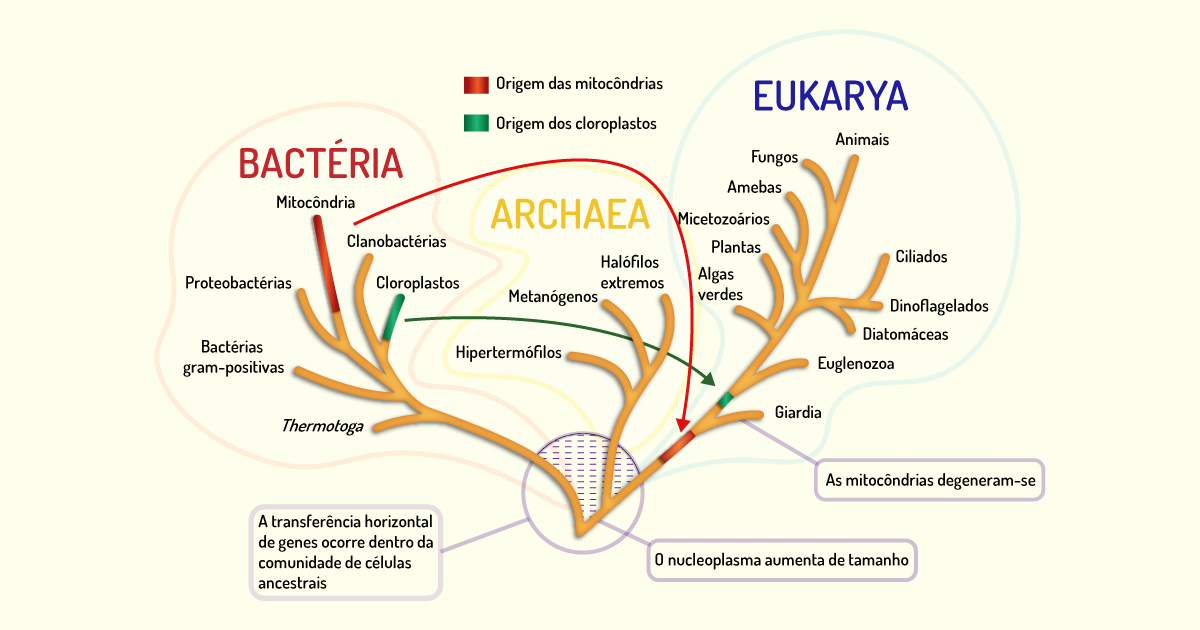
A árvore filogenética é uma ilustração gráfica que representa as relações entre os organismos que estão em estudo e é composta por nós e ramos. Os nós representam o ponto em que o ancestral divergiu em duas novas entidades, sendo que estas passaram a acumular diferenças durante sua evolução de forma independente. As pontas dos ramos também são chamadas de nós e representam linhagens individuais que existem atualmente. Já os ramos representam dois fatores: a ordem de descendência e a ancestralidade dos nós, sendo que o comprimento do ramo representa a quantidade de alterações que aconteceram ao longo do tempo (MADIGAN et al., 2016).
Nesta hierarquia filogenética, os organismos estão agrupados conforme propriedades comuns, isso sinaliza que um grupo de organismos teve sua origem a partir de um ancestral comum e que cada espécie mantém características de seu ancestral. Parte da informação utilizada para classificar os organismos é obtida por meio de fósseis. A maioria dos micro-organismos, porém, não é facilmente fossilizada. Dessa forma, outros indícios precisam ser utilizados para que a filogenia dos procariotos seja definida. Para isso, semelhanças nos genomas são utilizadas para classificar os micro-organismos em táxons e, também, para determinar uma cronologia para o surgimento desses organismos (TORTORA; FUNKE; CASE, 2017).
Na árvore filogenética (Figura 1.4), os organismos estão classificados de acordo com características similares. Além disso, esses organismos recebem um nome científico, composto por dois nomes (binômio). Esses nomes se referem ao gênero e ao epíteto específico e devem ser escritos sublinhados ou em itálico. O primeiro nome corresponde ao gênero e deve começar com letra maiúscula. O segundo nome corresponde à espécie, deve ser escrito com letras minúsculas e, geralmente, é um adjetivo que qualifica o gênero (TORTORA; FUNKE; CASE, 2017).
Esse esquema de classificação baseado em características similares auxilia na identificação dos organismos. No entanto os micro-organismos não são identificados necessariamente pelos mesmos métodos que os classificaram. Protozoários, fungos e vermes podem ser identificados com o auxílio de um microscópio. Grande parte dos micro-organismos, porém, não possui características distintas ou, ainda, possui uma grande variedade de morfologias. Diante disso, os microbiologistas desenvolveram uma grande variedade de procedimentos e critérios que auxiliam na identificação dos micro-organismos. Esses procedimentos são baseados na análise de diferenças na constituição da parede celular, na morfologia, na coloração diferencial, nas características metabólicas, nos testes bioquímicos etc. (TORTORA; FUNKE; CASE, 2017). Destacaremos, aqui, as técnicas de coloração, particularmente, a coloração de Gram.
Diversos compostos são utilizados para corar os micro-organismos. Os modos de ação de cada solução corante variam; alguns corantes se ligam a compostos químicos específicos presentes nas células, outros alteram a coloração na presença de reações químicas, dentre outros. Por meio das técnicas de coloração, os microbiologistas podem visualizar as estruturas dos micro-organismos, o que ajuda na identificação e na separação de micro-organismos similares (PELCZAR; CHAN; KRIEG, 2011).
As etapas para corar um micro-organismo para exame ao microscópio consistem em:
As técnicas de coloração podem ser coloração simples ou coloração diferencial. Na coloração simples, uma única solução corante é utilizada para corar o micro-organismo. O esfregaço, após ser fixado, é coberto com a solução corante por um período de tempo, em seguida, é lavado com água e seco. Geralmente, nessa técnica, as células são coradas de forma uniforme. Já na coloração diferencial, mais de uma solução corante é utilizada no processo, no qual as soluções são adicionadas uma após a outra. Assim, é possível visualizar determinadas partes da célula ou diferenças entre as células (PELCZAR; CHAN; KRIEG, 2011).
A técnica de coloração diferencial mais importante e utilizada é a coloração de Gram. Nessa técnica, o esfregaço é tratado com várias soluções na seguinte ordem: corante púrpura cristal violeta, solução de iodeto (mordente – solução que fixa o corante no interior da célula), álcool (descoloração – tem por finalidade remover o corante de determinadas bactérias) e, por fim, um corante vermelho, geralmente, safranina. Todas as etapas da técnica estão ilustradas na Figura 1.5) (PELCZAR; CHAN; KRIEG, 2011).
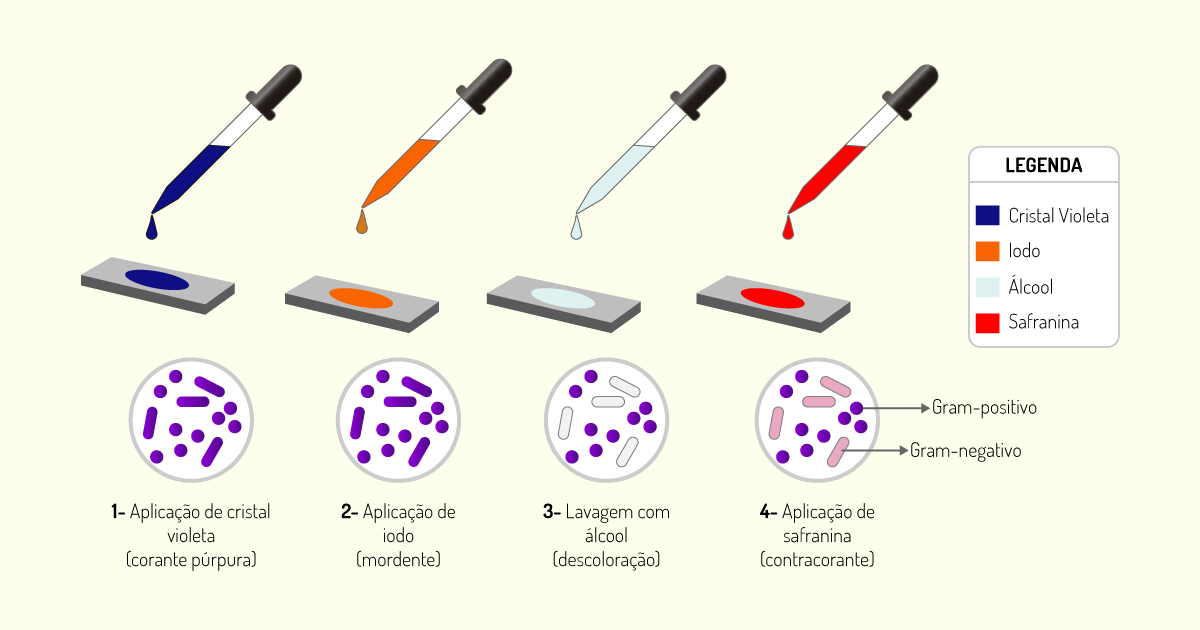
Ao final do processo, algumas bactérias apresentam-se coradas de violeta-escuro e são chamadas de Gram-positivas; outras apresentam coloração avermelhada por ação da safranina e são chamadas de Gram-negativas (Figura 1.6). As bactérias Gram-negativas apresentam coloração vermelha, pois tiveram o corante violeta removido quando foram tratadas com álcool e ficaram coradas apenas pelo último corante adicionado (PELCZAR; CHAN; KRIEG, 2011).
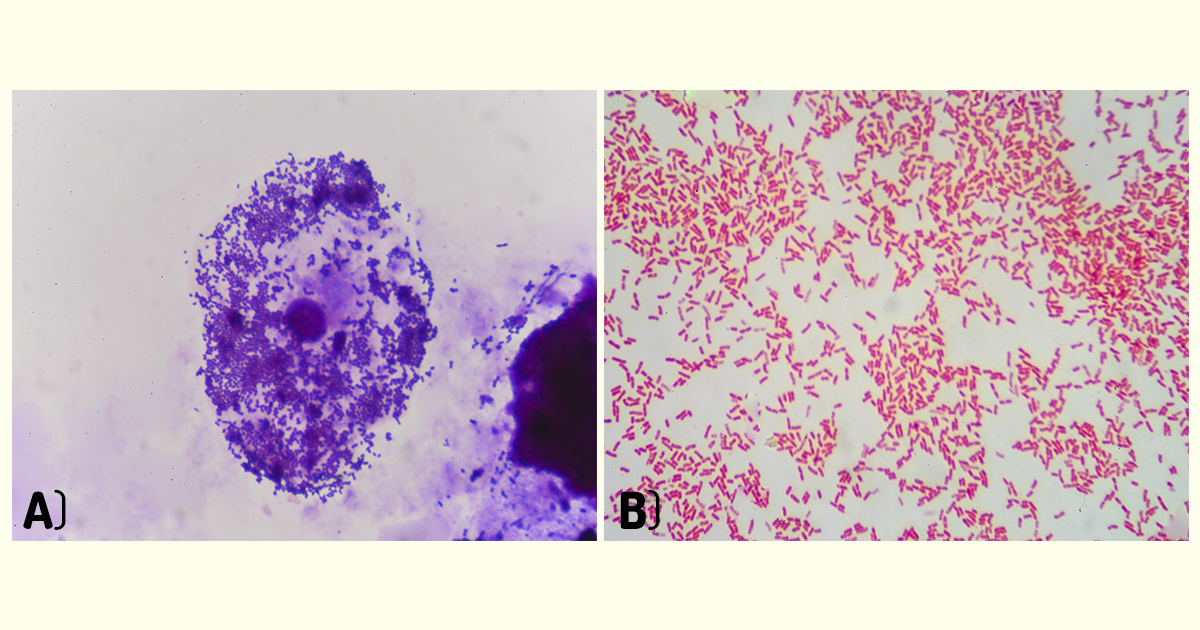
O princípio da técnica baseia-se na diferença de constituição estrutural das paredes das células das bactérias. Bactérias Gram-positivas possuem uma camada de peptideoglicano mais espessa, quando comparada com bactérias Gram-negativas. Já as bactérias Gram-negativas possuem, principalmente, lipopolissacarídeos em sua parede celular. No procedimento de coloração, as bactérias Gram-positivas formam um complexo cristal violeta-iodo, que não é facilmente removido quando tratado com álcool. Já as bactérias Gram-negativas não retêm o corante violeta, pois o tratamento com o álcool rompe com a camada externa de lipopolissacarídeos e remove o complexo cristal violeta-iodo; em seguida, são reveladas com o contracorante safranina, apresentando a coloração vermelha. Embora as bactérias Gram-positivas e Gram-negativas possam absorver o corante vermelho, esse corante é mascarado nas bactérias Gram-positivas pelo corante violeta. Dessa forma, por meio dessa técnica de coloração, esses dois tipos de células (Gram-positiva e Gram-negativa) podem ser diferenciados ao microscópio (PELCZAR; CHAN; KRIEG, 2011; MADIGAN et al., 2016; TORTORA; FUNKE; CASE, 2017).
A coloração de Gram é um procedimento de rotina e muito útil para distinguir diferentes tipos bacterianos. Normalmente, quando é necessário caracterizar um novo tipo de bactéria, isso é feito, inicialmente, determinando se a bactéria é Gram-positiva ou Gram-negativa (MADIGAN et al., 2016). Por exemplo, bactérias Gram-positivas encontradas em um esfregaço sanguíneo, arranjadas em cadeias curtas, sugerem uma infecção causada por estreptococos. Essa informação é útil para selecionar o tratamento para o paciente. As bactérias Gram-positivas são mais vulneráveis à penicilina. Já as bactérias Gram-negativas são mais resistentes, pois os antibióticos não conseguem penetrar na camada de lipopolissacarídeo (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
Caro(a) aluno(a), neste tópico, examinamos a classificação dos organismos e destacamos a técnica de coloração de Gram, que auxiliam na identificação dos micro-organismos. Nos próximos tópicos, estudaremos com mais detalhes essas classificações, discutindo as propriedades dos principais micro-organismos de cada grupo. Vamos iniciar a nossa discussão sobre a diversidade dos micro-organismos, enfocando os procariotos. Segundo o Bergey’s Manual of Systematic Bacteriology (principal publicação dedicada à classificação bacteriológica), os procariotos estão divididos em dois domínios: Bacteria e Archaea. Escritos com letras minúsculas bactérias e arqueas são, respectivamente, os organismos que pertencem a esses grupos. Lembrando que os domínios dividem-se em filos; os filos, em classes; as classes, em ordens; as ordens, em famílias; as famílias, em gêneros; e os gêneros, em espécies (MADIGAN et al., 2016).
A descoberta dos micro-organismos está associada à invenção do microscópio. A comunidade científica questionava a descoberta desses organismos desde a origem deles. Considerando os eventos, os experimentos e as teorias que marcaram o desenvolvimento da microbiologia, analise as afirmativas a seguir e assinale a alternativa correta.
Segundo a teoria da geração espontânea, uma célula só poderia ter origem a partir de outra célula pré-existente.
Alternativa incorreta, pois a geração espontânea acredita que alguns organismos podem surgir a partir de matéria inanimada, como moscas que “surgem” a partir de estrume . A teoria da biogênese defendia que uma célula só poderia ter origem a partir de outra célula pré-existente.
Os processos desenvolvidos por Pasteur foram aperfeiçoados e, hoje, formam a base das técnicas de assepsia.
Alternativa correta. Os experimentos que Pasteur utilizou para refutar a teoria da geração espontânea foram refinados e formam a base das técnicas de assepsia que eliminam e previnem a contaminação por micro-organismos indesejáveis.
Segundo a teoria da biogênese, as doenças estavam, de alguma maneira, relacionadas com os micro-organismos.
Alternativa incorreta, pois a teoria da biogênese definia que uma célula só poderia ter origem a partir de outra célula pré-existente. Segundo a teoria do germe e da doença, os germes estavam relacionados, de alguma maneira, com as doenças. A primeira evidência de que bactérias podem causar doenças foi de Robert Koch, que estudou o antraz, uma doença que estava causando a morte dos rebanhos de gado e ovelhas da Europa.
Os micro-organismos, por não serem organismos complexos, são todos unicelulares e estão agrupados apenas no domínio Bacteria.
Alternativa incorreta. Os micro-organismos podem ser unicelulares com as bactérias, no entanto também existem organismos multicelulares, como fungos. Os micro-organismos estão distribuídos nos três domínio Bacteria, Archea e Eucarya.
Na técnica de coloração de Gram, as bactérias Gram-positivas apresentam coloração vermelha, e as bactérias Gram-negativas apresentam coloração violeta.
Alternativa incorreta, pois, ao final do processo de coloração de Gram, algumas bactérias apresentam-se coradas de violeta-escuro e são chamadas de Gram-positivas, e outras apresentam coloração avermelhadas por ação da safranina e são chamadas de Gram-negativas. As bactérias Gram-negativas apresentam coloração vermelha, pois tiveram o corante violeta removido quando foram tratadas com álcool e ficaram coradas apenas pelo último corante adicionado. Essas diferenças se dão em virtude das diferentes constituições da parede celular dessas bactérias.
Como dissemos anteriormente, os procariotos estão divididos em dois grupos: Bacteria e Archaea. A principal diferença entre esses dois grupos está relacionada com a constituição da parede celular. A parede celular dos organismos que pertencem ao domínio Bacteria é formada por peptidoglicanos; já a parede celular dos organismos que pertencem ao domínio Archaea é constituída por proteína e polissacarídeos. A partir de agora, examinaremos a morfologia desses dois grupos de procariotos (PELCZAR; CHAN; KRIEG, 2011).
Vimos, anteriormente, como é realizada e qual o princípio da coloração de Gram. Essa técnica auxilia na diferenciação entre dois grupos de bactérias: as Gram-positivas e as Gram-negativas. O domínio Bacteria pode ser dividido em três grupos principais: as bactérias Gram-negativas, as bactérias Gram-positivas e os micoplasmas (que não formam parede celular). A seguir, estudaremos a diversidade morfológica desses três principais grupos de Bacteria (PELCZAR; CHAN; KRIEG, 2011).
As bactérias Gram-negativas, além do seu caráter Gram-negativo, possuem uma complexa parede celular externa, que cobre uma fina camada de peptidoglicano. Muitas das bactérias mais estudadas fazem parte desse grupo. As bactérias Gram-negativas são divididas em subgrupos, de acordo com características morfológicas e fisiológicas, como necessidade de oxigênio e motilidade. Veremos, a seguir, esses subgrupos (PELCZAR; CHAN; KRIEG, 2011).
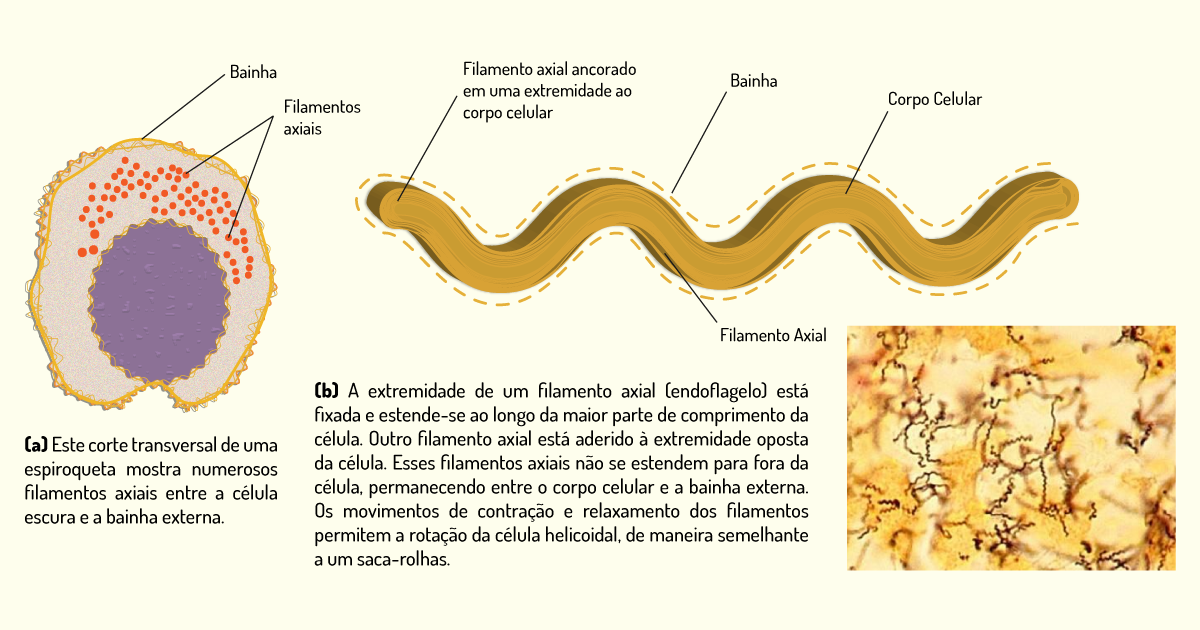
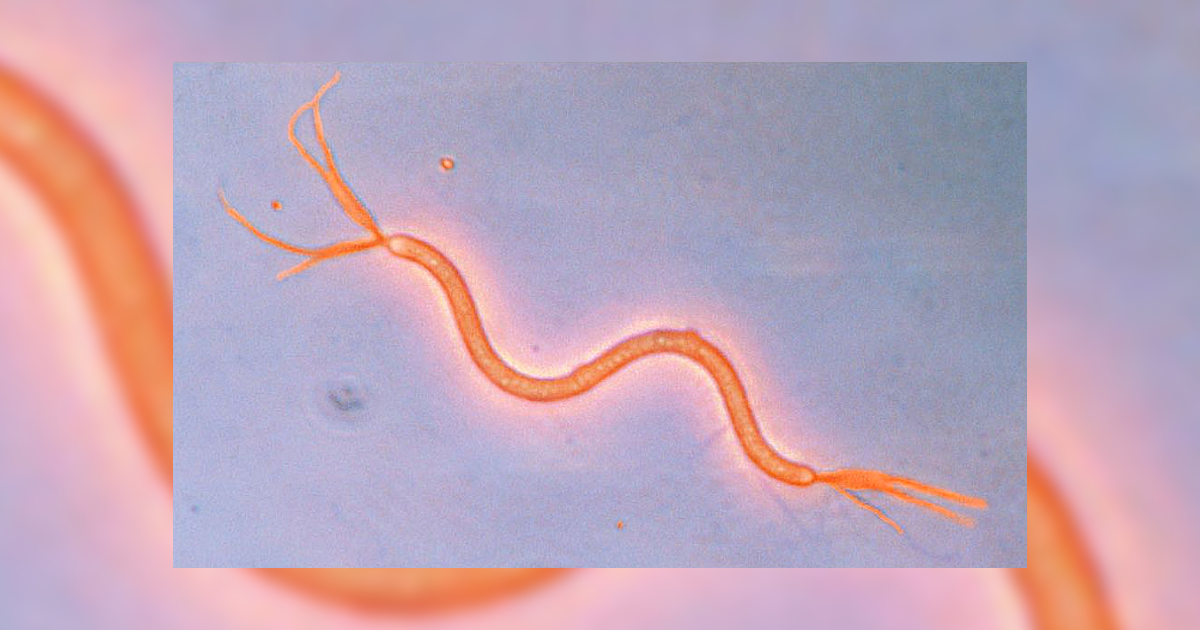
Espiroquetas – espiroquetas apresentam morfologia helicoidal espiralada, são flexíveis e móveis. Diferem das outras bactérias Gram-negativas flageladas com relação à localização do flagelo. Ao invés de estender seus flagelos para fora da célula, os flagelos ficam sob a membrana externa, ou seja, ficam localizados no espaço entre a membrana externa e o corpo da célula. Por esse motivo, recebem o nome de flagelo periplásmico. Sua movimentação se dá pela rotação de seu filamento axial, fazendo com que a célula gire na direção oposta, como uma saca-rolhas. Alguns espiroquetas são responsáveis por doenças como a sífilis, causada pelo Treponema pallidum (Figura 1.7), e algumas espécies de Borrelia, responsáveis por causar a doença de Lyne e a febre recorrente (PELCZAR; CHAN; KRIEG, 2011).
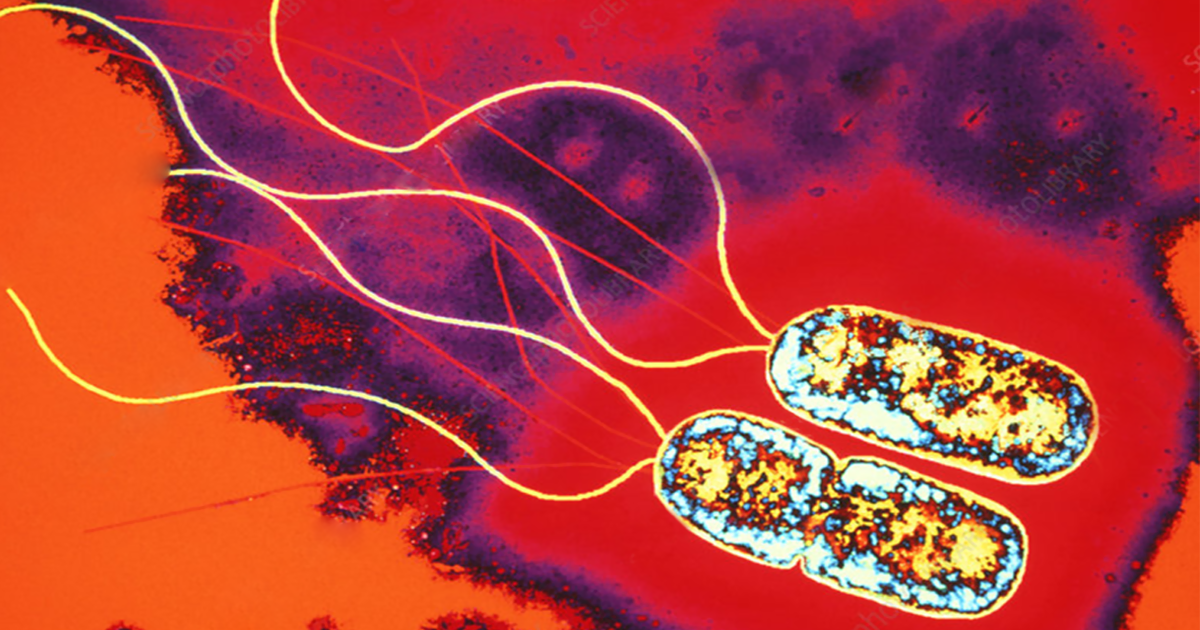
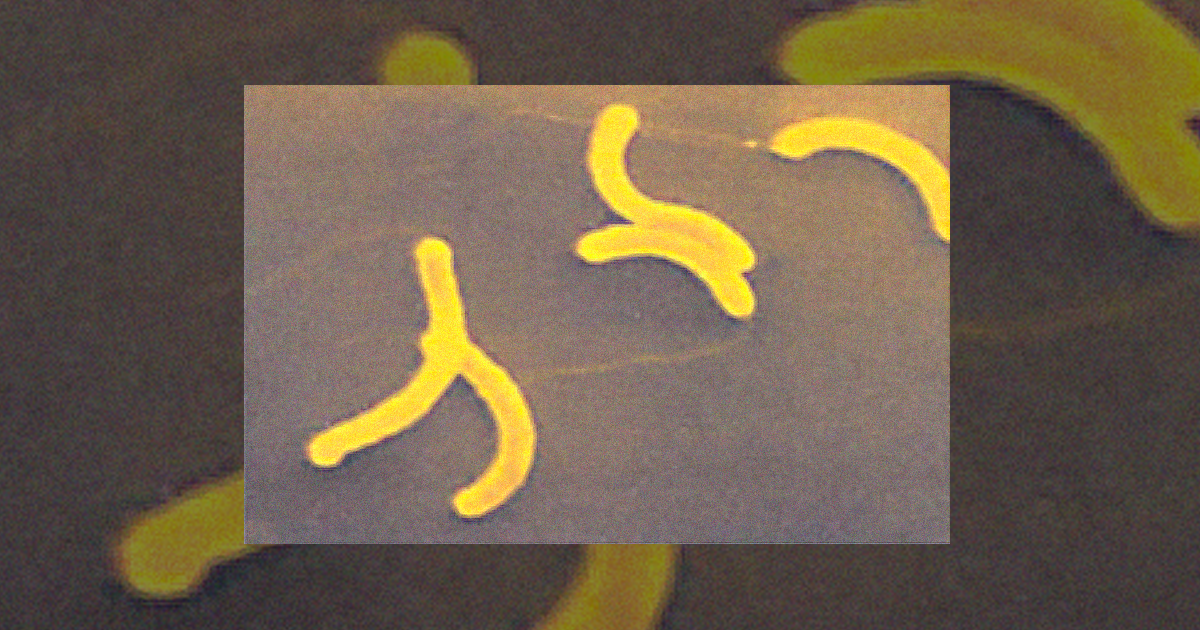
Espirilo – embora as bactérias desse grupo sejam semelhantes aos espiroquetas, os espirilos são proteobactérias rígidas, não flexíveis e espiraladas. Sua locomoção se dá por meio de flagelos polares (único ou múltiplo) localizados nas extremidades da célula. Podem ser aeróbios ou microaerofílicos, além de ter relação com plantas (simbiontes ou patógenos) ou animais (patógenos). Dentro desse grupo, podemos citar como exemplos os gêneros Spirillum (Figura 1.8) e Azospirillum. Cabe aqui ressaltar a importância do Azospirillum lipoferum, um organismo fixador de nitrogênio. Essa espécie chamou a atenção pelo fato de que participa de associações simbióticas com plantas como trigo e milho (MADIGAN et al., 2016; PELCZAR; CHAN; KRIEG, 2011).
Cocos e Bacilos Aeróbios – muitas bactérias Gram-negativas aeróbias que apresentam morfologia de bacilos ou bastonetes são muito comuns em solo e outros ambientes naturais. Um exemplo comum são as bactérias do gênero Pseudomonas (Figura 1.9). Essas bactérias são conhecidas pela sua capacidade de degradar diversos compostos orgânicos e utilizá-los como fonte de energia (PELCZAR; CHAN; KRIEG, 2011).
Nesse grupo de bactérias com essa morfologia, podemos destacar ainda a Azotobacter e o Rhizobium, que têm a habilidade de fixar o nitrogênio. As azotobactérias fixam nitrogênio do solo e são de vida livre, já os rizóbios fixam nitrogênio no interior de raízes de plantas como a soja (PELCZAR; CHAN; KRIEG, 2011).
Como exemplo de coco Gram-negativo aeróbico podemos citar a Neisseria gonorrhoeae (Figura 1.10), que, geralmente, ocorre como diplococo, habita as mucosas de mamíferos e é o agente causador da gonorreia (TORTORA; FUNKE; CASE, 2017).
Bacilos facultativos – nesse tipo de morfologia, estão inclusos os bacilos retos e encurvados. Por serem facultativos, podem se desenvolver aeróbica ou anaerobicamente. O exemplo mais comum desse grupo são os bacilos retos da família Enterobacteriaceae. Essas bactérias são denominadas entéricas e estão presentes no trato intestinal do homem e de animais de sangue quente. Elas possuem exigências nutricionais simples e fermentam açúcares, o que leva à produção de diferentes produtos finais (MADIGAN et al., 2016). O exemplo mais comum é a bactéria Escherichia coli, provavelmente, o organismo mais conhecido da microbiologia, considerada uma importante ferramenta na pesquisa biológica básica. A presença dessa bactéria em alimentos ou na água indica contaminação fecal. A E. coli pode estar envolvida com funções nutricionais no trato intestinal, pela síntese de vitaminas, como a vitamina K. Pelo fato de ser um organismo facultativo, provavelmente, auxilia no consumo de oxigênio e faz com que o intestino grosso tenha baixa oxigenação (anóxico) (MADIGAN et al., 2016). Outros patógenos humanos também são entéricos, como a Salmonella, que é o agente causador da febre tifoide e da gastroenterite, e as espécies de Shigella, agente causador da disenteria bacilar. Além das bactérias entéricas, podemos citar como anaeróbio facultativo o Vibrio cholerae (Figura 1.11), agente causador da cólera (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
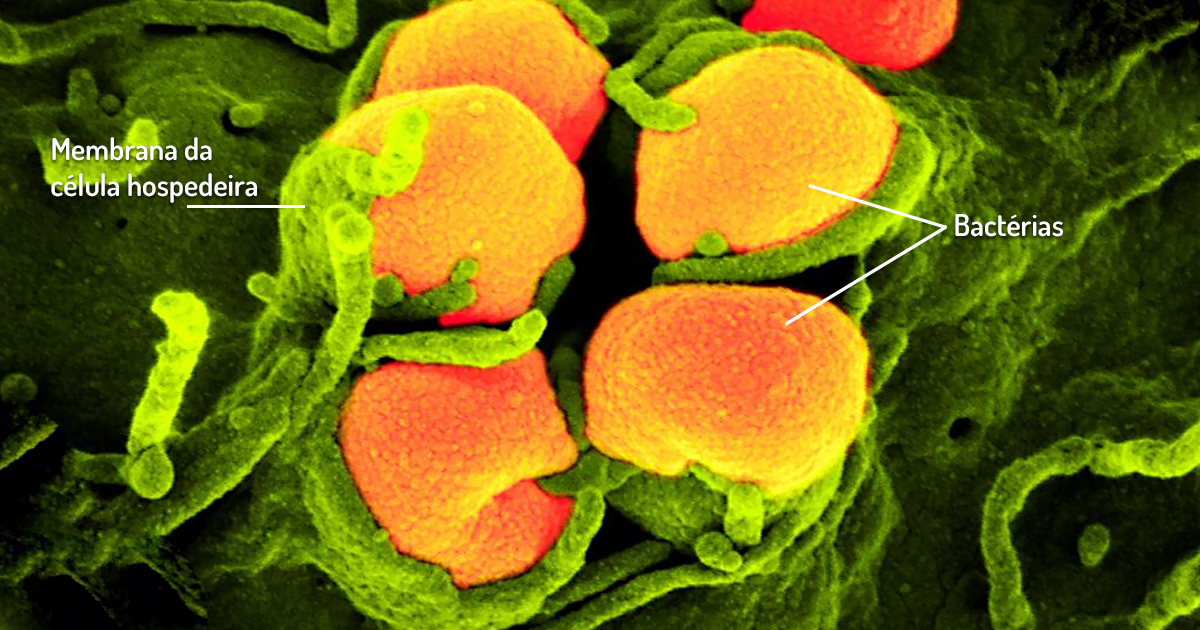
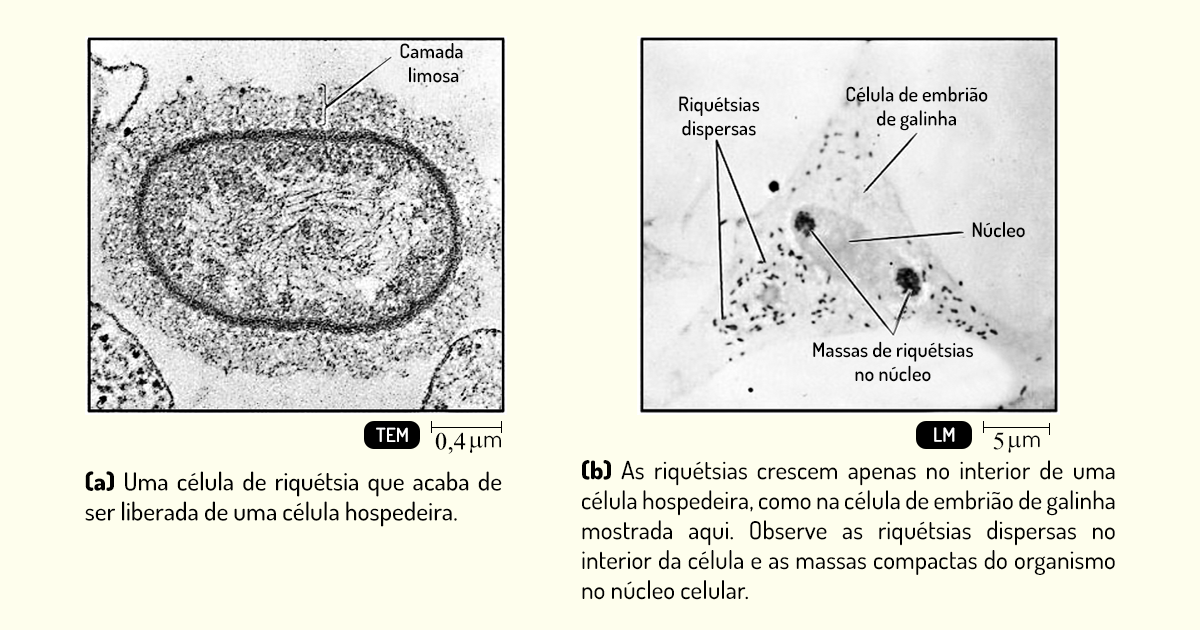
Riquétsias e Clamídias – tanto as riquétsias quanto as clamídias são parasitas intracelulares obrigatórias. As riquétsias (Figura 1.12) são parasitas com morfologia de bastonete ou cocobacilo que se desenvolvem na superfície ou no interior de células de artrópodes. Como são parasitas celulares obrigatórios, ainda não foram cultivadas sem as células hospedeiras (MADIGAN et al., 2016). São responsáveis por causar doenças em humanos, como a febre das Montanhas Rochosas (R. rickettsii). Muitas dessas doenças podem ser transmitidas para o homem por meio de piolhos e carrapatos que estejam infectados (PELCZAR; CHAN; KRIEG, 2011).
Já as clamídias são bactérias cocoides, não possuem peptidoglicano em sua parede celular, além de não necessitarem de insetos para a transmissão. Essas bactérias são transmissíveis para o homem por meio das vias respiratórias e do contato interpessoal. As espécies patogênicas dessas bactérias são C. psittaci, causadora da psitacose em pássaros, e C. trachomatis, agente causador da tracoma, que, no passado, foi uma das principais causas de cegueira no mundo; além da Chlamydophila pneumoniae, agente causador de uma forma branda de pneumonia que ocorre em jovens adultos (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
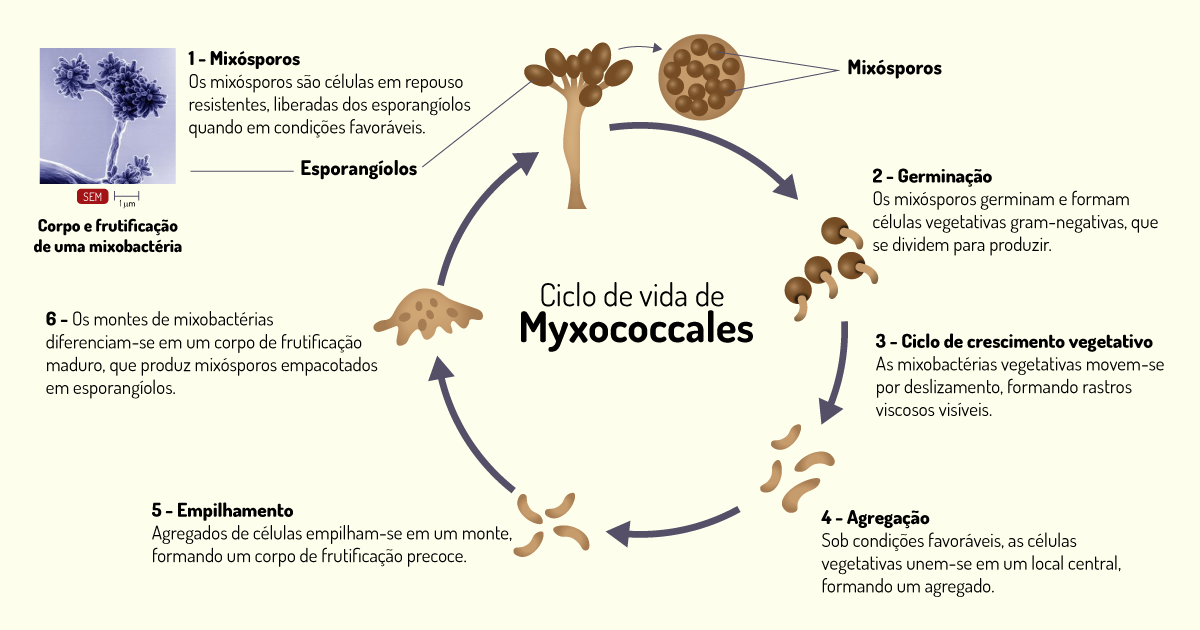
Bactérias deslizantes – existem dois tipos de bactérias deslizantes: as que formam corpos de frutificação (estruturas produtoras de esporos) e as que não formam essas estruturas. As bactérias que formam os corpos de frutificação são bastonetes curtos e flexíveis, não possuem flagelos, porém se movem por deslizamento sobre superfícies, deixando rastro de limo. Assim, essas bactérias são denominadas myxobactérias, que significa “bactéria limosa”. Essas bactérias, à medida que se movem, destroem enzimaticamente e digerem outras bactérias que encontram pelo caminho, utilizando-as como fonte de nutrientes.
Quando estão no mesmo estágio de crescimento, as células se agregam e se diferenciam, formando corpos de frutificação, que contêm células em repouso, chamadas de mixósporos. Essa diferenciação é induzida por baixas quantidades de nutrientes. Assim que níveis de nutrientes passam a ser apropriados, há a formação de novas células vegetativas deslizantes a partir dos mixósporos. Você pode observar o ciclo de vida de uma mixobactéria na Figura 1.13. Essas bactérias são aeróbias e podem ser encontradas na superfície do solo, por exemplo, a matéria orgânica em decomposição (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
As bactérias deslizantes que não formam frutificação são bastonetes ou são filamentosas aeróbias e não formam os mixósporos. Uma característica interessante dessas bactérias é que elas são capazes de degradar alguns polímeros naturais, como a celulose, a queratina e, até mesmo, o ágar (PELCZAR; CHAN; KRIEG, 2011).
Prezado(a) aluno(a), examinamos, até agora, a morfologia das bactérias Gram-negativas. Passaremos, a partir do próximo tópico, a estudar a morfologia das bactérias Gram-positivas.
Vimos, nos tópicos anteriores, que a parede celular das bactérias Gram-positivas e Gram-negativas possuem diferenças em sua constituição. As bactérias Gram-positivas têm uma parede celular mais espessa e não possuem uma membrana externa. Essas bactérias são divididas de acordo com suas características morfológicas e bioquímicas.
Cocos – as bactérias Gram-positivas com essa morfologia, ou seja, com formato esférico, são divididas em grupos, de acordo com o arranjo celular e o tipo de metabolismo. O gênero Micrococcus, por exemplo, está arranjado de forma irregular ou em grupos de quatro. Talvez os mais conhecidos sejam os Staphylococcus e os Streptococcus. Os Staphylococcus são facultativos quanto à necessidade de oxigênio e são encontrados na pele e na membrana de humanos e animais de sangue quente. A espécie patogênica mais comum é o Staphylococcus aureus (Figura 1.14), agente causador de várias doenças, como síndrome do choque tóxico, intoxicações alimentares e infecções pós-operatórias (PELCZAR; CHAN; KRIEG, 2011).
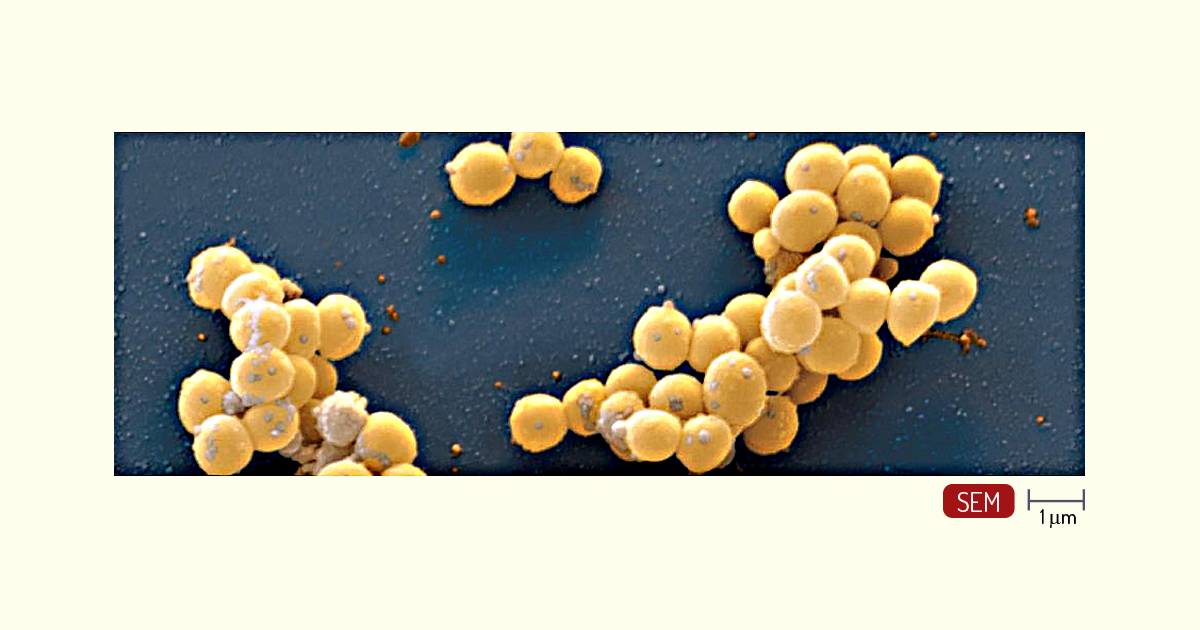
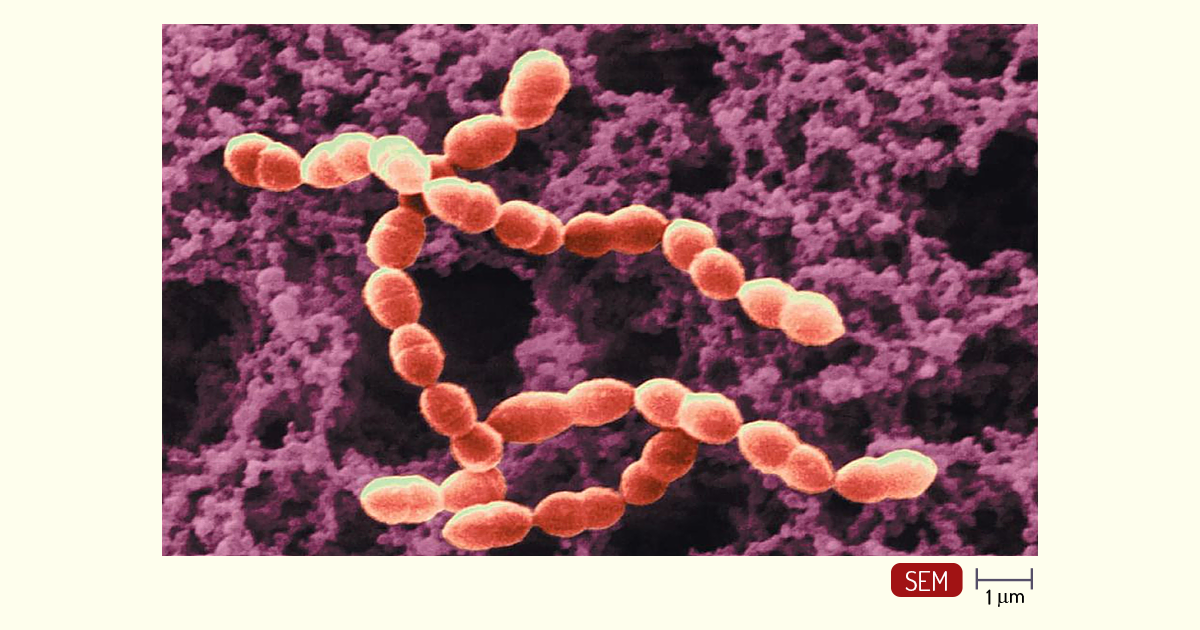
Os Streptococcus (Figura 1.15), diferentemente, são arranjados aos pares ou em cadeias. São responsáveis por causar várias doenças, dentre elas, febre reumática (S. pyogenes), pneumonia bacteriana no homem (S. pneumoniae) etc. Dentre os mecanismos que contribuem para a patogenicidade, está a produção de substâncias extracelulares que destroem as células fagocíticas, que são as células responsáveis por eliminar organismos e substâncias estranhas ao corpo (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
Bactérias esporuladas – algumas bactérias formam estruturas chamadas de endósporos, que são resistentes a uma grande variedade de agentes, como desinfetantes, calor e radiação. A grande maioria das bactérias que formam esporos tem morfologia de bastonetes, com exceção do Sporosarcina, que é cocoide. Os gêneros Bacillus e Sporosarcina são aeróbios ou facultativos. Muitas espécies são saprófitas, ou seja, se alimentam de matéria orgânica em decomposição, e são encontradas em vários ambientes, como solo, água doce ou mar. O Clostridium (Figura 1.16) é um exemplo de bactéria esporulada anaeróbia. As espécies Clostridium botulinum e Clostridium tetani produzem neurotoxinas, que são responsáveis pelo botulismo e pelo tétano, respectivamente (PELCZAR; CHAN; KRIEG, 2011).
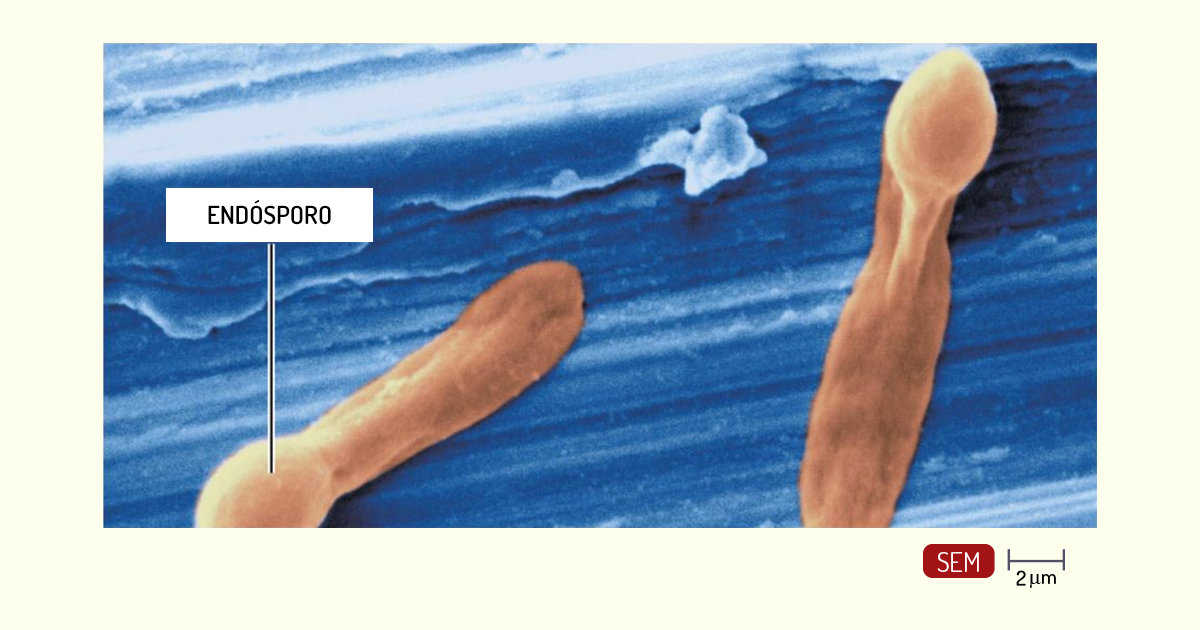
Caro(a) aluno(a), acabamos de estudar as bactérias esporuladas. Vimos que algumas são responsáveis por causar doenças muito sérias, como o botulismo e o tétano. No entanto existem algumas dessas bactérias esporuladas que são utilizadas como inseticidas, pois são capazes de infectar insetos. Um exemplo é a bactéria esporuladas Gram-positiva Bacillus thuringiensis, que produz cristais de uma toxina protéica (toxina Bt) quando esporula. Dessa forma, quando os insetos ingerem a bactéria, esses cristais se dissolvem no trato intestinal dos insetos, causando sua morte. Preparações comerciais com os endósporos e os cristais dessa bactéria são vendidas como produtos para jardinagem, para ser pulverizadas sobre as plantas. É importante lembrar de que esses cristais não são tóxicos para os humanos e animais e, por serem uma proteína, são degradados e não se acumulam no ambiente.
Fonte: Pelczar, Chan e Krieg (2011).

Bacilos regulares - alguns bastonetes não esporulados possuem a morfologia uniforme, ou seja, não apresentam saliências, ramificações ou outras variações de forma. Quanto às necessidades de oxigênio, são aeróbios, facultativos ou anaeróbios. Podemos citar como exemplo para esse grupo os Lactobacillus, que são bactérias presentes nos processos fermentativos de produtos animais e vegetais, ou, ainda, presentes nas mucosas de humanos, como boca, trato gastrointestinal e vagina. Geralmente, não são considerados patogênicos, são utilizados na indústria para a fabricação de produtos lácteos, como iogurtes e queijos. No entanto, algumas espécies são patogênicas, como a Listeria monocytogenes, que pode ser adquirida de queijo ou leite não pasteurizado adequadamente e pode ser o agente causador de abortos espontâneos e natimortos (PELCZAR; CHAN; KRIEG, 2011).
Bacilos irregulares – além dos bacilos regulares, existem bactérias não esporuladas irregulares. Essas bactérias são bastonetes levemente curvados, apresentam saliências, possuem ramificações e outros desvios na morfologia. Podemos citar como exemplo a Arthrobacter que, inicialmente, se desenvolve como bastonetes regulares, tornando-se cocos; posteriormente, entretanto, quando são inoculadas em meio fresco, desenvolvem-se novamente como bastonetes. Esse ciclo de vida é denominado ciclo coco-bacilo (PELCZAR; CHAN; KRIEG, 2011).
Actinomicetos – essas bactérias têm a características de formar micélios, que são emaranhados de filamentos denominados hifas. São encontrados no solo e atuam na degradação de restos de plantas e animais. Um importante gênero dentro desse grupo de bactérias é o Streptomyces, que é utilizado para a produção de antibióticos, como a tetraciclina e a estreptomicina (PELCZAR; CHAN; KRIEG, 2011).
Prezado(a) aluno(a), neste tópico, dedicamo-nos a estudar a morfologia de procariotos. A princípio, mencionamos que as bactérias podem ser divididas de acordo com as características de sua parede celular em três grupos: Gram-negativas, Gram-positivas e micoplasma. Já estudamos os dois primeiros grupos. A partir de agora, estudaremos o grupo dos micoplasmas.
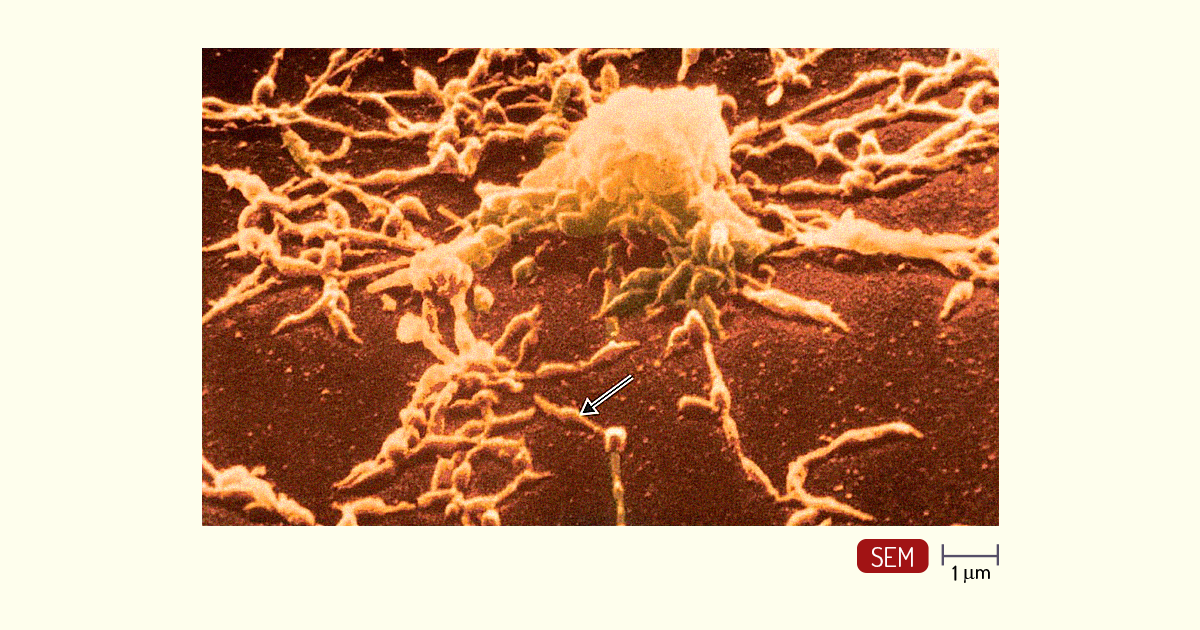
Os micoplasmas têm como característica a incapacidade de formar parede celular, sendo assim, possuem apenas uma membrana citoplasmática que o envolve externamente. Dessa forma, essas bactérias coram-se como Gram-negativas. Devido à ausência da parede celular, essas bactérias possuem algumas propriedades, como:
Muitos micoplasmas possuem em sua membrana citoplasmática colesterol, uma característica de células eucarióticas. Dessa forma, o meio de cultura para essas bactérias contém soro sanguíneo, com o objetivo de fornecer o colesterol necessário para seu desenvolvimento. Eles crescem 5 m nesse tipo de meio, por isso se tornaram um problema recorrente de contaminação de culturas celulares em laboratório. Um exemplo de micoplasma é a espécie patogênica Mycoplasma pneumoniae (Figura 1. 17), agente causador de um tipo de pneumonia (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017). Essas bactérias dependem de seu hospedeiro para sobreviver, assim, são incapazes de viver como organismos de vida livre (TORTORA; FUNKE; CASE, 2017).
Prezado(a) aluno(a), finalizamos nosso estudo sobre a morfologia das bactérias. O próximo assunto será dedicado à diversidade morfológica das arqueas.
Como mencionamos no início desta unidade, as arqueas, também chamadas de arqueobactérias, são organismos que vivem em ambientes extremos, como ambientes com elevadas temperaturas (piscinas termais) ou lagos com elevadas concentrações salinas. As arqueas são divididas em três grupos principais: halófilos, acidófilos e termófilos. Os dois primeiros grupos possuem parede celular, e os termoplasmas têm como característica ausência de parede celular (TORTORA; FUNKE; CASE, 2017).
Halófilos – são denominadas halobactérias (“bactérias do sal”), pois necessitam de ambientes com quantidades superiores a 25 % de NaCl para terem um bom desenvolvimento. As halobactérias não se desenvolvem em soluções com concentrações de sal abaixo de 15 %, nem mesmo a água do mar, com, aproximadamente, 3% de NaCl, é salgada o suficiente para o desenvolvimento dessa bactérias. Dessa forma, as halobactérias se desenvolvem apenas em ambientes que não são propícios para outros organismos, como o Mar Morto. Devido à presença de caratenoides, essas bactérias apresentam coloração entre o vermelho e o laranja, entretanto não possuem clorofila. Ao que parece, esses caratenoides protegem essas bactérias contra os efeitos prejudiciais da luz solar (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
Acidófilos – esse grupo de arqueas é encontrado em fontes de água quente ácida. Podem crescer entre temperatura de 50 ºC e 87 ºC e preferem condições ácidas, sendo que algumas espécies podem crescer em pH baixos. Arqueas do gênero Sulfolobus, cujo pH ideal é 2, oxidam o enxofre ou os compostos orgânicos como açúcares para obtenção de energia (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
Termófilos – os termoplasmas são semelhantes aos micoplasmas, devido à ausência de parede celular e por possuírem uma membrana citoplasmática que os envolve externamente; porém diferem deles por serem capazes de crescer em ambientes com altas temperaturas, podendo crescer em temperaturas a partir de 80 ºC. O recorde de temperatura para crescimento é de 121 ºC, registrado por arquea termófila, que vive próxima a fontes termais a 2000 m de profundidade no oceano (PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017).
Caro(a) aluno(a), finalizamos nosso segundo tópico da Unidade I, em que nos dedicamos a estudar a diversidade morfológica dos procariotos. Vimos os diferentes exemplos e morfologias de organismos importantes em cada grupo, tanto do domínio Bateria quanto Archeae. Nos próximos tópicos, dedicaremo-nos a estudar os outros grupos de micro-organismo. A seguir, falaremos sobre os fungos.
Os procariotos são um grupo de organismos que têm como característica serem unicelulares. Eles são divididos em dois grupos Bacteria e Archaea. Considerando as características desses organismos, analise as afirmativas a seguir e julgue a alternativa correta:
Os três principais grupos de Bacteria são bactérias Gram-positivas, bactérias Gram-negativas e bactérias halofílicas.
Alternativa incorreta, pois os três principais grupos de Bacteria são as bactérias Gram-positivas, as Gram-negativas e o micoplasma. As halofílicas pertencem a Archea.
Os três principais grupos de Archaea são termófilos, halófilos e acidófilos.
Alternativa correta, as bactérias que pertencem ao grupo das Archeae vivem em ambientes extremos, como ambientes com elevadas temperaturas (termófilos), ambientes ácidos (acidófilos) e ambientes com altas concentrações de sal (halófilos).
Os parasitas são bactérias que retiram nutrientes de meio orgânico inanimado.
Alternativa incorreta, bactérias que retiram nutrientes de matéria orgânica inanimada, como material em decomposição, são chamadas de saprófitas. Parasitas são organismos que necessitam de um hospedeiro vivo para obter seus nutrientes.
Micoplasmas são bactérias desprovidas de parede celular que vivem em ambientes com elevadas temperaturas.
Alternativa incorreta, os micoplasmas, de fato, não possuem parede celular, entretanto não sobrevivem em ambientes com altas temperaturas. Os organismos que crescem em altas temperaturas e não possuem parede celular são os termófilos, que pertencem ao domínio Archeae.
Bacilos regulares têm como característica possuir uma morfologia uniforme e um ciclo de vida denominado ciclo coco-bacilo.
Alternativa incorreta, pois, no ciclo de vida coco-bacilo, a bactéria cresce como bastonetes, tornando-se, mais tarde, cocos e, se inoculada em meio fresco, torna-se bastonetes novamente. Há uma irregularidade na morfologia, sendo assim, esse ciclo de vida é característico de bacilos irregulares, e não de bacilos regulares, como expõe a alternativa.
Prezado(a) aluno(a), passaremos, a partir de agora, a estudar os micro-organismos eucarióticos. Assim como os micro-organismos procarióticos, os micro-organismos classificados como eucarióticos possuem ampla diversidade morfológica e processos celulares. São organismos fascinantes, devido ao seu complexo ciclo de vida e por sua aplicação econômica. Neste tópico, iremos nos dedicar a estudar, particularmente, os fungos.
Os fungos fazem parte do reino Fungi, são organismos eucarióticos e não fotossintetizantes. São reconhecidos três grupos de fungos: os bolores, as leveduras e os cogumelos. Com exceção das leveduras, os fungos são multicelulares, sendo que alguns são macroscópicos. Além disso, possuem parede celular e produzem esporos. Podem ser aeróbios ou anaeróbios facultativos, com poucas exceções conhecidas de anaeróbios. Os fungos também são quimio-heterotróficos, necessitando de fontes de carbono. A grande maioria dos fungos não é patogênica. Esses organismos são benéficos, pois têm papel importante na decomposição de matéria vegetal morta, o que leva à reciclagem de elementos vitais PELCZAR; CHAN; KRIEG, 2011; TORTORA; FUNKE; CASE, 2017). A seguir, estudaremos as características que auxiliam na identificação dos fungos.
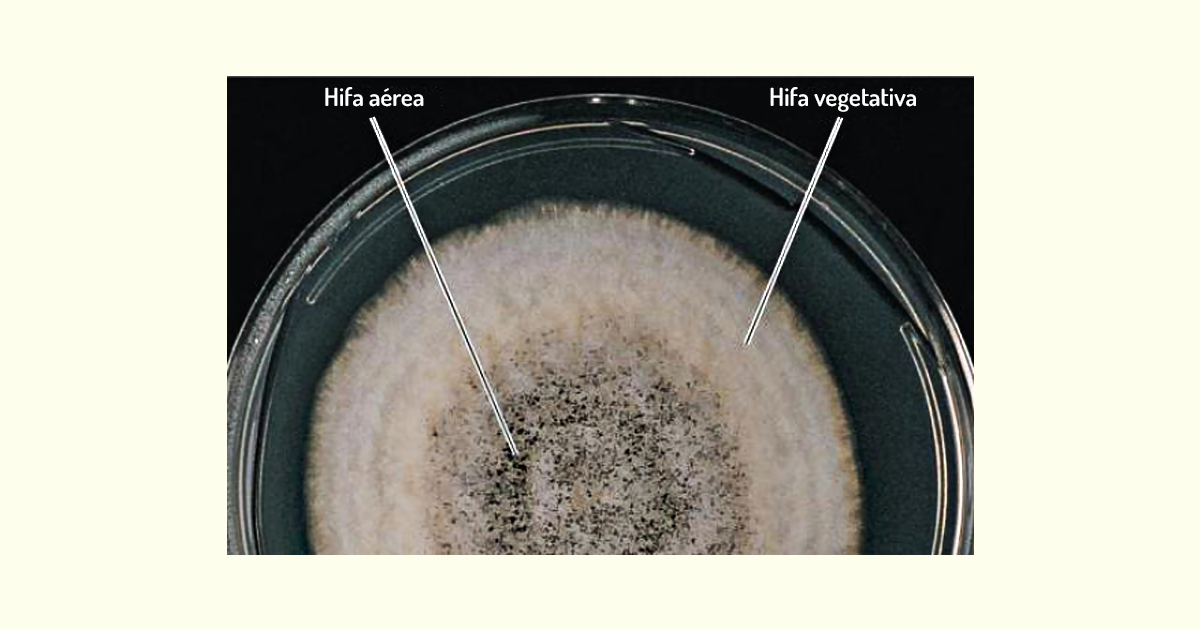
Bolores – os bolores são fungos filamentosos que consistem em longos filamentos de células denominados hifas. Em grande parte dos bolores, as hifas contêm septos, que são como paredes cruzadas que dividem a hifas em unidades distintas, semelhantes às células. Essas hifas são denominadas hifas septadas (Figura 1.17a). Entretanto, em alguns fungos, as hifas não possuem septos; essas células são, portanto, contínuas e apresentam vários núcleos, sendo chamadas de hifas cenocíticas (Figura 1.17b). As hifas septadas, no entanto, possuem canais entre os septos, o que torna contínuo o citoplasma das unidades adjacentes. Além disso, as hifas têm porções com funções distintas. A porção responsável por obter nutrientes é denominada hifa vegetativa, e a porção relacionada à reprodução chama-se hifa reprodutiva ou aérea. Na Figura 1.17b, há o crescimento de uma hifa a partir de um esporo; note que ela cresce por alongamento (TORTORA; FUNKE; CASE, 2017).
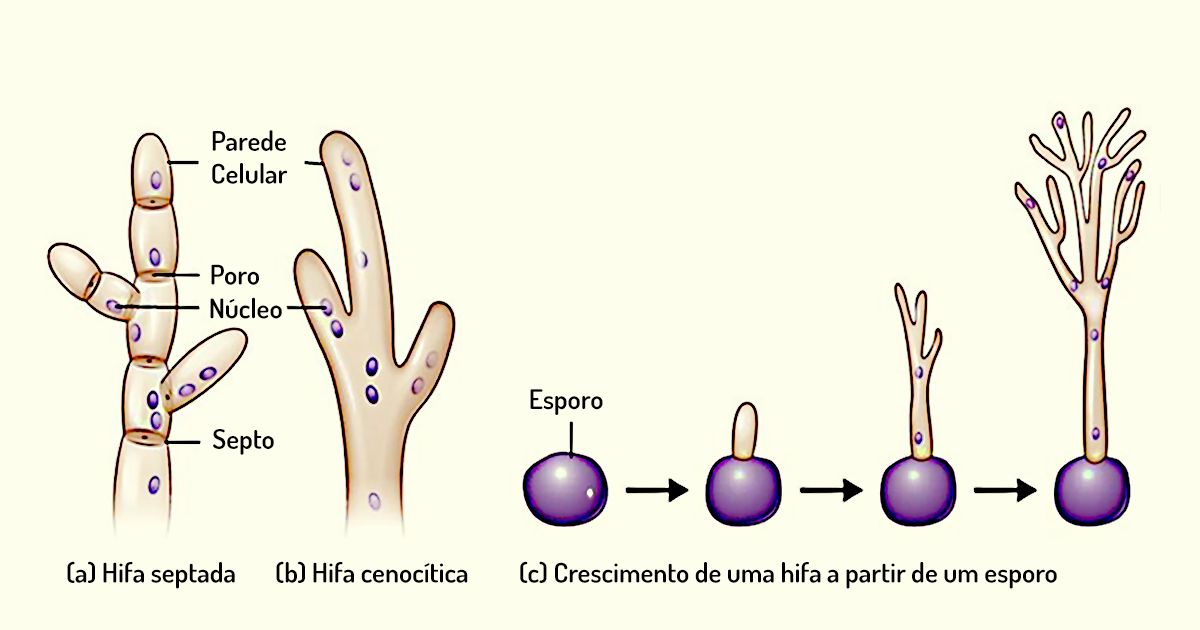
Muitas vezes, as hifas aéreas atuam como suporte dos esporos reprodutivos. Em condições ambientais favoráveis, as hifas formam micélios que correspondem a uma massa filamentosa, que pode ser observada a olho nu (Figura 1.18).
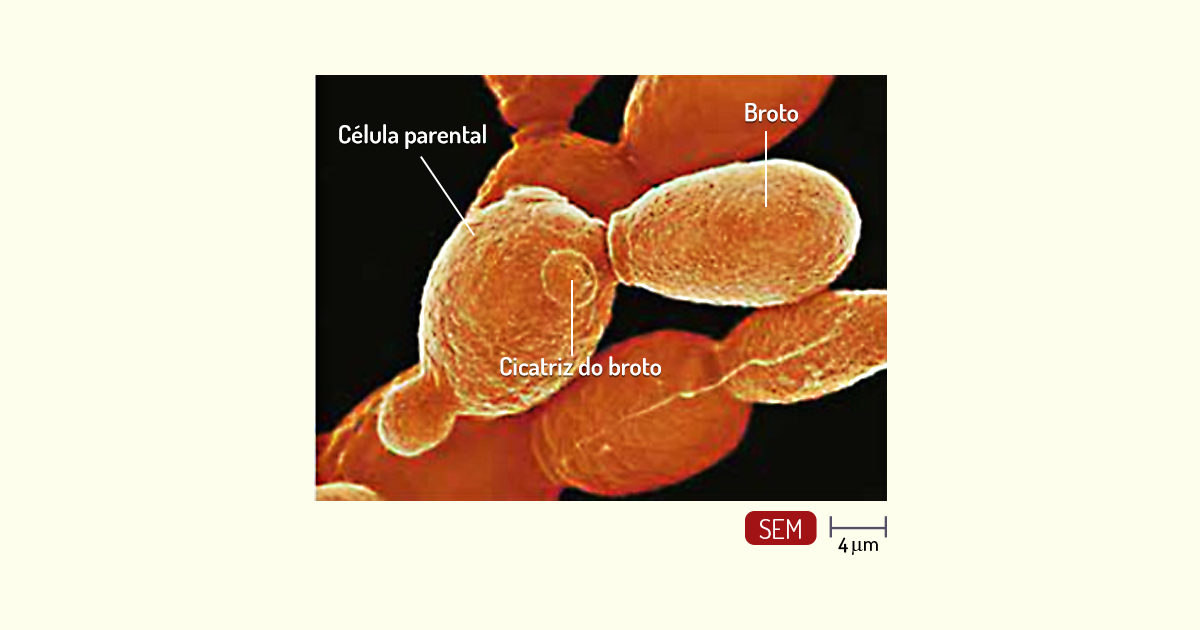
Leveduras – as leveduras são fungos unicelulares não filamentosos. Quanto à morfologia, as células de levedura são esféricas ou ovais e, geralmente, a divisão celular ocorre por brotamento. Nesse processo (Figura 1.19), uma nova célula é formada como uma protuberância (broto) na superfície da célula parental. O broto se alonga, o núcleo da célula parental sobre divisão, e um dos núcleos se direciona para o broto. Então, o broto se separa da célula parental (MADIGAN et al., 2010; TORTORA; FUNKE; CASE, 2017).
Embora grande parte das leveduras dividam-se como células individuais, algumas leveduras, sob certas condições, produzem brotos que não se separam, formando pequenas cadeias, denominadas pseudo-hifa. A Candida albicans, capaz de causar infecções orais, pulmonares e vaginais, necessita dessa fase com pseudo-hifas para invadir tecidos profundos. No processo de brotamento, as células se dividem de maneira desigual, diferente das células que se dividem por fissão, gerando duas células iguais. Nesse processo, a célula parental alonga-se, o núcleo sofre divisão e, então, as células filhas são produzidas (MADIGAN et al., 2016; TORTORA; FUNKE; CASE, 2017).
As leveduras podem ser anaeróbios facultativos; quando há oxigênio, elas são aeróbias e metabolizam carboidratos, formando dióxido de carbono. Na ausência de oxigênio, as leveduras fermentam carboidratos, produzindo etanol e dióxido de carbono. Um exemplo é a Saccharomyces, amplamente utilizada na indústria, que produz etanol nas bebidas fermentadas e dióxido de carbono nos pães, fazendo a massa crescer (TORTORA; FUNKE; CASE, 2017).
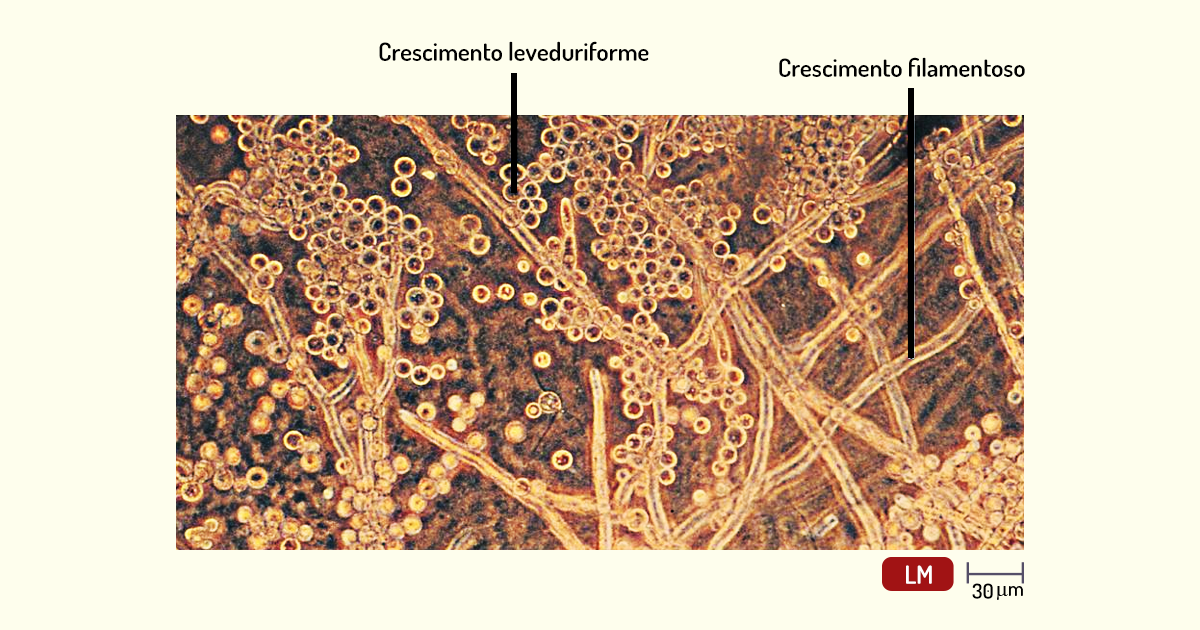
Fungos dimórficos – alguns fungos exibem dimorfismo, ou seja, apresentam duas formas de crescimento; podem crescer tanto como levedura como fungo filamentoso. Esse dimorfismo em fungos patogênicos depende de fatores como a temperatura e a concentração de \(CO_2\). Na Figura 1.20, podemos observar o fungo Mucor indicus, em que seu dimorfismo depende da concentração de \(CO_2\). Nos locais onde o \(CO_2\) do metabolismo se acumulou, o fungo apresenta crescimento filamentoso e, na superfície do ágar, esse fungo apresenta crescimento leveduriforme (TORTORA; FUNKE; CASE, 2017).
Agora, prezado(a) aluno(a), listaremos alguns fungos contaminantes de alimentos e de culturas em laboratórios.
Zigomiceto – são fungos filamentosos que apresentam hifas cenocíticas. Podemos citar o Rhizopus stolonifer, um fungo bem conhecido como bolor do pão. Seus esporos são assexuados e chamados de esporangiósporos, que ficam dentro do esporângio. Quando os esporangiósporos se dispersam por locais com condições adequadas, podem germinar e dar origem a um novo talo de fungo.
Ascomycota – no grupo dos ascomicetos, incluem-se fungos que têm hifas septadas e algumas leveduras; seus esporos, normalmente, são conídios e assexuados. O termo “conídio” significa “pó”, assim, esses esporos são facilmente liberados e flutuam como poeira no ar. Quando os núcleos de duas células se fundem, formam um ascósporo. Esses esporos são produzidos em estruturas denominadas sacos (TORTORA; FUNKE; CASE, 2017).
Basidiomycota – os basidiomicetos são fungos com hifas septadas. Esse grupo inclui os fungos que produzem corpo de frutificação, os cogumelos. Os esporos, ou seja, os basidiósporos são formados externamente, em estruturas denominadas basídios, em que são formados quatro basidiósporos por basídio. Alguns fungos desse grupo produzem conidiósporos assexuados (TORTORA; FUNKE; CASE, 2017).
Quanto à importância econômica desses organismos, os fungos já são utilizados há muitos anos. A Saccharomyces cerevisiae, por exemplo, é amplamente empregada na indústria alimentícia para a produção de pães e vinhos. O Aspergillus niger, por sua vez, é utilizado para a produção de ácido cítrico para alimentos. Em relação aos fármacos, o fungo Taxomyces é usado na produção de taxol, um fármaco antitumoral. Outra utilização dos fungos é na área de controle biológico, como o fungo Entomophaga que, em 1990, proliferou-se, de forma inesperada, nos EUA, mas que acabou eliminando mariposas que estavam prejudicando árvores (TORTORA; FUNKE; CASE, 2017).
Como você pode observar, caro(a) aluno(a), são inúmeras as aplicações dos fungos, desde a produção de alimentos até a eliminação de pragas. Nesse sentido, muitas pesquisas são realizadas com o objetivo de melhorar a utilização desses organismos tão fascinantes.
Embora os fungos estejam associados a doenças e a prejuízos, como a contaminação de alimentos, eles podem ser usados de forma benéfica, visto que possuem diversas aplicações. No texto “A importância dos fungos na biotecnologia”, as autoras Camila Joyce Alves da Silva e Diana Jussara do Nascimento Malta discutem, por exemplo, a utilização desses organismos para a produção de novos produtos biotecnológicos, e abordam o potencial dos fungos em relação à recuperação ambiental. Você pode ler o texto, na íntegra, no link: periodicos.set.edu.br.

Os fungos são organismos que pertencem ao reino Fungi; são organismos eucarióticos e, com exceção das leveduras, são multicelulares. Considerando as características desses organismos, assinale a alternativa correta.
Os bolores são organismos que, para se dividirem, realizam divisão por brotamento ou fissão, dividindo-se simetricamente.
Alternativa incorreta, pois esses processos de divisão celular são característicos das leveduras.
Os fungos possuem inúmeras aplicações e podem ser utilizados na fabricação de alimentos e fármacos.
A Saccharomyces cerevisiae, por exemplo, é um fungo utilizado na fabricação de pães e vinhos. Por sua vez, o fungo Taxomyces é utilizado na produção do antitumoral taxol.
As leveduras têm hifas que podem ser septadas, ou seja, contêm estruturas denominadas septos, que dividem a hifa em unidades que lembram células.
Alternativa incorreta, pois as leveduras, morfologicamente, não têm hifas. Essa característica está ligada aos bolores.
As leveduras, quando estão em condições ambientais favoráveis, produzem uma massa filamentosa denominada micélio.
Alternativa incorreta, pois são os bolores que, em condições ambientais favoráveis, produzem micélio.
O grupo dos basidiomicetos inclui leveduras e fungos com hifas septadas, cujos esporos são conídios e assexuados.
Alternativa incorreta, pois os indivíduos citados na alternativa pertencem ao grupo dos ascomicetos.
Prezado(a) aluno(a), para finalizar esta unidade, apresentaremos os conceitos relacionados às algas, aos protozoários e aos vírus, abordando a morfologia, o crescimento e a classificação desses organismos.
“As algas são organismos eucariotos fotoautotróficos, relativamente simples, que não possuem os tecidos (raízes, caules e folhas) típicos de plantas” (TORTORA; FUNKE; CASE, 2017, p. 332). De acordo com essa definição, quando se trata das algas, não estamos nos referindo a um grupo taxonômico particular, ou seja, os organismos chamados de algas não estão classificados todos juntos, em um mesmo reino, como ocorre com os fungos, que você estudou anteriormente.
Nesse sentido, o termo “alga” é utilizado como referência aos organismos fotoautotróficos que não apresentam raízes e caules como os das plantas. As algas já foram consideradas plantas em algum momento da história, mas, por não apresentarem os embriões das plantas verdadeiras, deixaram de ser. Desse modo, com o passar dos anos e o avanço científico na área da análise de DNA, as algas foram reclassificadas em diferentes reinos (TORTORA; FUNKE; CASE, 2017).
A diversidade de formas é bastante grande entre as algas: algumas são unicelulares e outras são chamadas de filamentosas, porque formam cadeias de células; algumas são bem visíveis, como é o caso das algas marrons que, eventualmente, chegam às praias, outras apenas formam uma espécie de espuma em poças de água ou manchas verdes sobre o solo. A maioria das espécies é aquática ou vive em locais úmidos, porque as algas precisam da água para o suporte físico, a reprodução e a difusão de nutrientes (TORTORA; FUNKE; CASE, 2017).
As algas apresentam uma estrutura chamada de talo, que corresponde ao corpo da alga. Nas algas grandes, multicelulares, conhecidas como algas marinhas, o talo consiste em apressórios ramificados (que servem para fixá-las em rochas), além de estruturas chamadas de hastes cauliformes que, geralmente, são ocas, e lâminas, que se parecem com folhas (TORTORA; FUNKE; CASE, 2017).
As células que recobrem o talo são fotossintetizantes, e não existe um tecido condutor, como xilema e floema: os nutrientes são absorvidos por toda a superfície da alga. A água circundante é quem sustenta o talo da alga, pois as hastes não são lignificadas nem lenhosas, diferente do caule das plantas. Ademais, algumas algas são sustentadas pelo pneumatocisto, uma vesícula flutuante, cheia de gás (TORTORA; FUNKE; CASE, 2017).
As algas podem se reproduzir tanto sexuadamente como assexuadamente. A reprodução assexuada ocorre por fragmentação, no caso de algas multicelulares e filamentosas, sendo que cada pedaço tem a capacidade para formar um novo talo ou filamento. No caso das algas unicelulares, ocorre a mitose, que é um processo de divisão celular no qual a célula duplica seu DNA e, posteriormente, divide-se. Independente do processo, essas formas de reprodução assexuada originam clones, ou seja, algas geneticamente iguais à alga que as originou. A reprodução sexuada, por sua vez, envolve a formação e a fertilização dos gametas para a formação do zigoto.
Algumas espécies apresentam várias gerações seguidas de reprodução assexuada, mas pode acontecer de, devido à alguma condição do meio, a mesma espécie se reproduzir de forma sexuada. Além disso, outras espécies apresentam alternância de gerações, pois a prole resultante da reprodução sexuada se reproduz assexuadamente, e a geração seguinte volta a apresentar reprodução sexuada (TORTORA; FUNKE; CASE, 2017).
A classificação das algas se baseia em semelhanças e diferenças de estrutura, pigmentos, e em algumas outras características morfológicas, além de sequências de rRNA (RNA ribossômico, uma região do genoma bastante usada em estudos taxonômicos, a partir da comparação da sequência entre os indivíduos). As algas pertencem ao Domínio Eukaria e, a seguir, estão descritos os principais grupos de algas.
Algas marrons – são algas marinhas (mais encontradas em regiões costeiras), multicelulares, macroscópicas, que chegam a atingir 50 m de comprimento. As paredes celulares delas contêm celulose e ácido algínico (a algina, um espessante utilizado em muitos alimentos, é extraída da parede celular dessas algas). Esse tipo de alga pertence ao reino Chromalveolata, filo Phaeophyta (TORTORA; FUNKE; CASE, 2017).
Algas vermelhas – vivem em regiões mais profundas do oceano, pois seus pigmentos vermelhos permitem a absorção da luz azul que chega até essas regiões. A maioria delas é multicelular e a parede celular é composta por celulose. Algumas espécies possuem talos que formam uma crosta sobre conchas e rochas. Esse tipo de alga pertence ao reino Archaeplastida e ao filo Rhodophyta. O ágar, utilizado em meios de cultura para o cultivo de micro-organismos, é extraído dessas algas (TORTORA; FUNKE; CASE, 2017).
Algas verdes – em sua maioria, são microscópicas. Essas algas pertencem ao filo Chlorophyta, do reino Viridiplantae, e podem ser unicelulares ou multicelulares. Assim como as plantas, a parede celular das algas verdes é composta de celulose, há as clorofilas “a” e “b” e elas armazenam o amido. Acredita-se que as plantas terrestres surgiram a partir das algas verdes (TORTORA; FUNKE; CASE, 2017).
Diatomáceas – algas unicelulares ou filamentosas, incluídas no reino Chromalveolata, no filo Bacillariophyta. A parede celular nessas algas é complexa e contém pectina e sílica. Além disso, as diatomáceas produzem algumas toxinas que podem causar intoxicação alimentar (TORTORA; FUNKE; CASE, 2017).
Dinoflagelados – são as algas unicelulares flutuantes, conhecidas como plâncton. Elas pertencem ao reino Chromalveolata e ao filo Dinoflagellata. A celulose presente na membrana confere rigidez a essas algas. Ademais, alguns dinoflagelados produzem neurotoxinas; e grandes concentrações de Alexandrium (uma espécie de dinoflagelado) causam o fenômeno conhecido como maré vermelha, quando o oceano se tinge com uma forte coloração avermelhada (TORTORA; FUNKE; CASE, 2017).
Mofos aquáticos – são também chamados de bolores aquáticos e classificados no reino Chromalveolata e filo Oomycota. São organismos multicelulares e a maioria é de decompositores, formando massas sobre algas e animais mortos, geralmente, em água doce. São organismos que já foram classificados como fungos, devido à semelhança com os fungos zigomicetos (produção de esporos em um esporângio, porém, ao contrário dos esporos do fungo, os esporos dos oomicetos têm dois flagelos) (TORTORA; FUNKE; CASE, 2017).
Assim como as plantas são importantes nas cadeias alimentares terrestres, as algas desempenham esse papel nos ambientes aquáticos: elas realizam a fixação do CO2 em moléculas orgânicas, que serão consumidas pelos demais organismos. Além disso, o O2 também é liberado na fotossíntese. As algas que vivem nas partes mais superficiais dos corpos de água (mar, rio, lago etc.) são chamadas de planctônicas (os primeiros metros).
Você já parou para pensar na quantidade de água que existe no nosso planeta? 75% do território da Terra é coberto por água. A estimativa é que 80% do O2 da Terra seja produzido por algas planctônicas. Esses dados mostram a importância da preservação desses corpos de água, pois uma grande parte dos seres vivos, incluindo você , depende do oxigênio (TORTORA; FUNKE; CASE, 2017).
Os protozoários compõem um grupo bastante grande e variado dentre os micro-organismos. Apesar de o termo “protozoário” significar “primeiro animal”, em razão da semelhança entre a nutrição dos protozoários e dos animais, esses organismos são bastante diferentes dos animais. Isso porque existem protozoários que realizam fotossíntese, além de várias espécies terem um ciclo de vida complexo, no qual podem existir mudanças de hospedeiros para as espécies parasitas (várias doenças que atingem o homem são causadas por protozoários).
Assim como as algas, a reprodução dos protozoários pode ser sexuada ou assexuada. A reprodução assexuada ocorre por fissão, na qual a célula se divide, originando duas células iguais; por brotamento, quando a célula forma um broto, que cresce e se desprende, originando um novo organismo; ou por esquizogonia, uma fissão múltipla, na qual o núcleo divide-se várias vezes e uma pequena porção do citoplasma se concentra ao redor de cada núcleo e, no fim, a célula se separa em várias células-filhas.
Por outro lado, a reprodução sexuada pode ocorrer por conjugação ou pela produção de gametas. Neste momento, caro(a) aluno(a), é importante que você não confunda esse processo com o bacteriano de conjugação: você aprenderá sobre ele na Unidade II. No processo de conjugação entre os protozoários, duas células se fundem e seus núcleos haploides (denominados micronúcleos) também se fundem. Quando essas duas células parentais se separam, cada uma delas se torna uma célula fertilizada que, ao se dividir, produz células-filhas, com o DNA recombinado. Por sua vez, os gametas – células sexuais haploides que se fundem durante a reprodução para a formação de um zigoto diploide – são produzidos por algumas espécies de protozoários (TORTORA; FUNKE; CASE, 2017).
Quando as condições do ambiente se tornam adversas (ausência de alimento, umidade ou oxigênio, presença de compostos tóxicos e temperaturas inadequadas), alguns protozoários são capazes de produzir uma cápsula protetora, denominada cisto. O cisto permite que o organismo sobreviva em um ambiente inóspito ou que uma espécie de protozoário parasita possa sobreviver fora do seu hospedeiro (TORTORA; FUNKE; CASE, 2017).
A classificação das espécies de protozoários, ou seja, o enquadramento das espécies em categorias taxonômicas, como reino e filo, é realizado de acordo com a morfologia e com análises de DNA. Conforme definimos no início deste tópico, os protozoários formam um grupo bastante diversificado, logo, uma morfologia generalizada do grupo deixaria muito a desejar. Por isso, caro(a) aluno(a), você poderá conhecer as principais características morfológicas à medida que for estudando a classificação desses micro-organismos.
Além disso, é importante esclarecer que essas classificações que envolvem os seres vivos não são fixas. Isso porque, com o avanço dos métodos de análise, tanto moleculares quanto microscópicos, podem ocorrer alterações, com o objetivo de melhorar o sistema de classificação. Atualmente, os protozoários estão classificados dentro do Domínio Eukarya. A seguir, estão descritos alguns grupos de protozoários e suas principais características.
Reino Excavatal – composto por todos os eucariotos unicelulares que possuem uma cavidade alimentar em seu citoesqueleto. A maioria dos organismos é fusiforme e possui flagelos. O reino Excavatal é um super-reino que engloba dois filos: um é composto por micro-organismos que não possuem mitocôndrias e o outro é o filo Euglenozoa .
As espécies parasitas Giardia intestinalis, que vivem no intestino delgado de mamíferos em geral, e Trichomonas vaginalis, que pode ser encontrada na vagina e no trato urinário masculino, são exemplos de protozoários que não possuem mitocôndrias e, portanto, estão nesse super-reino.
Em relação ao filo Euglenozoa, os membros desse grupo têm as seguintes características em comum: mitocôndrias em forma de disco, ausência de reprodução sexuada e sequências comuns no DNA. Existem, no entanto, dois grupos de células flageladas nesse filo: euglenoides e hemoflagelados.
Os euglenoides são fotoautotróficos (por isso, costumam ser estudados junto com as algas), possuem uma membrana plasmática semirrígida, denominada película, e se movem por meio de um flagelo localizado na extremidade anterior. A maioria dos euglenoides também apresenta uma organela contendo carotenoides (chamada de estigma vermelho), que percebe a luz e dirige a célula na direção apropriada.
Os hemoflagelados são os protozoários transmissíveis por meio de picadas de insetos hematófagos e encontrados no sangue do hospedeiro picado. Geralmente, eles têm corpos longos e delgados, além de uma membrana ondulante. A doença conhecida no Brasil como doença de Chagas é causada por um protozoário desse grupo, o Trypanosoma cruzi, que, por sua vez, é transmitido por um inseto conhecido como “bicho barbeiro” (TORTORA; FUNKE; CASE, 2017).
Amebas – são protozoários que apresentam estruturas chamadas de pseudópodes ou “falsos pés”. Esses pseudópodes são projeções citoplasmáticas que servem para a movimentação da ameba. A única espécie de ameba patogênica encontrada no intestino dos humanos é a Entamoeba histolytica, que coloniza cerca de 10% da população. A transmissão de Entamoeba ocorre pela ingestão dos cistos que são excretados nas fezes das pessoas infectadas. Os gêneros Acanthamoeba e Balamuthia contêm espécies de ameba de vida livre, que vivem na água, e que não são transmissíveis entre os seres humanos (TORTORA; FUNKE; CASE, 2017).
Apicomplexa – os protozoários que compõem esse filo têm um conjunto de organelas especiais nas extremidades de suas células (ou nos ápices, e disso deriva o nome do grupo). Essas organelas possuem enzimas cuja função é penetrar os tecidos do hospedeiro. A forma madura desses protozoários é imóvel, e eles são parasitas intracelulares obrigatórios. O ciclo de vida nesses organismos é complexo, porque envolve a transmissão entre vários hospedeiros. O Plasmodium, causador da malária, é um exemplo desse filo (TORTORA; FUNKE; CASE, 2017).
Ciliados – os protozoários ciliados possuem os cílios, estruturas semelhantes aos flagelos, porém mais curtos e numerosos. Esses cílios estão envolvidos com a movimentação da célula e com o direcionamento de partículas alimentares para a boca. Existe apenas uma espécie de protozoário ciliado que parasita seres humanos: Balantidium coli. Esse protozoário causa um tipo de disenteria grave, e a transmissão ocorre quando o hospedeiro ingere cistos que entram no intestino delgado, desenvolvendo-se nele (TORTORA; FUNKE; CASE, 2017).
Os cientistas começaram a usar o termo “vírus”, que, de acordo com a sua origem no latim, significa “veneno”, por volta de 1930. Eles queriam descrever um tipo de agente infeccioso, uma partícula, menor do que tudo que era conhecido até então. Em 1935, o primeiro vírus foi isolado: ele infectava plantas de tabaco, causando uma doença, o que tornou possível os estudos químicos e estruturais sobre os vírus.
Nesse contexto, uma questão persiste até os dias de hoje: os vírus são organismos vivos ou não? Os vírus não podem ser considerados organismos vivos, pois não apresentam características básicas e essenciais à vida, como todos os outros organismos, visto que não têm atividade metabólica e são parasitas celulares obrigatórios, ou seja, necessitam de um hospedeiro.
Entretanto os vírus podem ser considerados seres vivos, pois, uma vez no interior da célula hospedeira, são capazes de replicar seu ácido nucleico viral, infectar outros organismos e causar doenças. Desse modo, “dependendo do ponto de vista, um vírus pode ser considerado um agregado excepcionalmente complexo de elementos químicos ou um microrganismo vivo extraordinariamente simples” (TORTORA; FUNKE; CASE, 2017, p. 359).
Nesse sentido, a seguir, apresentaremos alguns aspectos importantes dos vírus e, assim, no fim da unidade, você mesmo, caro(a) aluno(a), pode formular sua própria resposta para a questão que gera tantas controvérsias.
O termo “vírion” é utilizado para descrever a partícula viral infecciosa completa, totalmente desenvolvida, que é formada por um ácido nucleico e um revestimento proteico. Você pode imaginar algo equivalente à membrana plasmática celular, mas cuja função é apenas proteger contra o ambiente externo. Os vírus são classificados de acordo com o ácido nucleico que possuem e por diferenças nas estruturas de seus envoltórios (TORTORA; FUNKE; CASE, 2017).
Enquanto, nas células de todos os outros organismos, o DNA corresponde ao material genético principal e o RNA desempenha uma função auxiliar, existem vírus cujo material genético é apenas o DNA, e outros que possuem apenas RNA. Outra diferença é que o DNA de todas as células consiste em uma molécula de fita dupla, e o RNA é, normalmente, fita simples. Nos vírus, além dessas duas formas, existem DNA fita simples e RNA fita dupla. Isso varia bastante e serve de suporte para a classificação dos vírus (TORTORA; FUNKE; CASE, 2017).
O revestimento proteico que protege o material genético do vírus é chamado de capsídeo, e as unidades proteicas que compõem o capsídeo são denominadas capsômeros. Geralmente, os capsômeros podem ser visualizados em microscópios eletrônicos. Em alguns vírus – chamados de vírus envelopados – existe uma estrutura denominada envelope, que envolve o capsídeo. A composição desse envelope, normalmente, consiste em uma combinação de lipídios, proteínas e carboidratos. Quando os vírus são liberados de uma célula hospedeira, por meio de um processo chamado de extrusão, a membrana plasmática dessa célula envolve a partícula viral e, nesses casos, o envelope viral passa a ser a membrana plasmática proveniente da célula (TORTORA; FUNKE; CASE, 2017).
Em alguns vírus, os envelopes podem apresentar espículas, que são complexos de carboidrato e proteína que, ao se projetarem para a superfície do envelope, auxiliam o vírus na ligação com a superfície de uma célula hospedeira. Essas espículas também são utilizadas para a identificação e a classificação dos vírus . Na Figura 1.21, há a morfologia de um vírus envelopado.
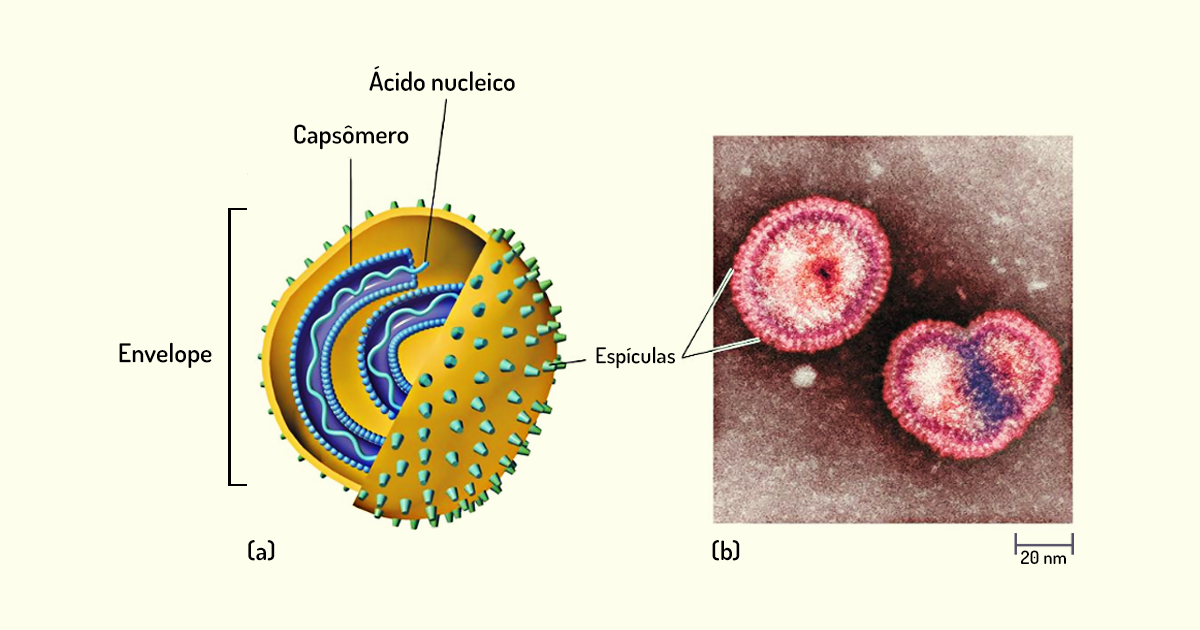
Os vírus também podem ser do tipo não envelopados, ou seja, o capsídeo não é envolvido por um envelope. Nesse caso, é o capsídeo que protege o ácido nucleico viral e se liga a partículas de células suscetíveis (TORTORA; FUNKE; CASE, 2017).
O material genético dos vírus contém apenas alguns genes necessários para a síntese de novos vírus, como os genes para as proteínas que formam o capsídeo e algumas enzimas envolvidas na replicação e na infecção viral. No entanto, para que essas proteínas e enzimas sejam sintetizadas e funcionem, o vírus precisa estar dentro da célula hospedeira, pois é ela que fornece energia, ribossomos e tudo o que o vírus precisa para se multiplicar. Portanto, o vírus não consegue se multiplicar se não estiver dentro de uma célula e utilizando sua maquinaria metabólica (TORTORA; FUNKE; CASE, 2017). Um único vírus pode originar até milhares de partículas virais iguais, mesmo considerando uma única célula hospedeira.
Nesse processo, esses vírus podem causar severos danos à célula hospedeira, levando-as à morte, mas alguns vírus não causam a destruição dessa célula, a qual, continuamente, produz novos vírus. Essa diferença se deve à existência de dois mecanismos alternativos de multiplicação: o ciclo lítico e o ciclo lisogênico. No ciclo lítico, os vírus provocam a lise e, portanto, a célula hospedeira morre, enquanto no ciclo lisogênico a célula hospedeira permanece viva. (TORTORA; FUNKE; CASE, 2017). Observe o esquema desses dois ciclos na Figura 1.22.
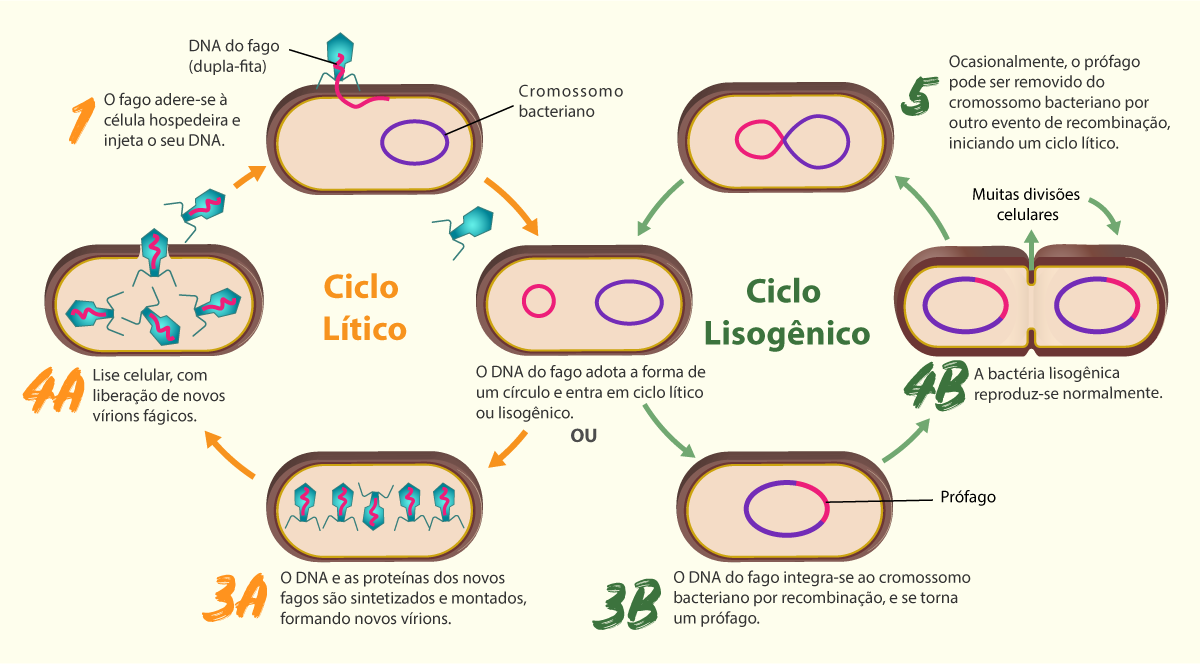
A seguir, apresentaremos a classificação dos vírus.
Caro(a) aluno(a), até aqui, você aprendeu que todos os organismos se enquadram em um dos três domínios: Archaea, Bacteria ou Eukarya, mas os vírus não são classificados como parte de qualquer um desses três domínios. Os vírus não são formados por células, ao contrário, apenas utilizam-nas para se multiplicarem. O nicho ecológico de um vírus, ou seja, o seu local mais íntimo de sobrevivência, é sua célula hospedeira específica, assim, pode ser que o vírus esteja mais relacionado ao seu hospedeiro do que a outros vírus (TORTORA; FUNKE; CASE, 2017).
O Comitê Internacional de Taxonomia de Vírus (International Committee on Taxonomy of Viruses) utiliza a expressão espécie viral para descrever uma população de vírus com características similares e que ocupa um nicho ecológico em particular (ou seja, um mesmo hospedeiro) (TORTORA; FUNKE; CASE, 2017).
Embora não estejam classificados como os demais micro-organismos, existe uma classificação dos vírus, também chamada de taxonomia viral, para ajudar a entender e a organizar novos vírus, quando descobertos. Por exemplo, vírus que são bastante parecidos entre si podem ser combatidos com um mesmo medicamento ou por meio do mesmo mecanismo de ação (TORTORA; FUNKE; CASE, 2017).
Assim como acontece com outros grupos, as análises das sequências do material genético dos vírus fornecem informações para que eles sejam agrupados em famílias, de acordo com o genoma de cada um e com a estrutura (envelopado ou não, DNA de fita simples ou dupla, RNA de fita simples ou dupla). Alguns exemplos de famílias virais são: Herpesviridae (causa doenças como herpes labial, catapora, herpes zóster e mononucleose infecciosa), Flaviviridae (febre amarela e dengue) e Retroviridae (vírus causador da AIDS) (TORTORA; FUNKE; CASE, 2017).
As algas e os protozoários são eucariotos, enquanto os vírus não podem ser classificados como procariotos ou eucariotos, pois não são formados por células. Além disso, não existe um reino específico, que reúna todas as espécies de algas ou protozoários, porque esses organismos são bastante diversificados. Com base nas diferenças e semelhanças entre algas, protozoários e vírus, assinale a alternativa correta.
As algas se multiplicam por um processo denominado esquizogonia, enquanto os protozoários realizam mitose.
A alternativa está incorreta, pois a esquizogonia é um processo de reprodução assexuada, que ocorre em protozoários. A mitose, por sua vez, refere-se a um processo realizado para a multiplicação das algas.
O principal material genético presente nas algas e nos protozoários é o DNA, enquanto os vírus podem conter tanto DNA quanto RNA.
A alternativa está correta, porque a informação genética dos vírus pode estar contida tanto em uma molécula de DNA quanto RNA. Por seu turno, o ácido nucleico principal, presente em todos os procariotos e eucariotos (como as algas e os protozoários), é o DNA.
Ao contrário das algas e dos vírus, os protozoários são organismos eucariotos fotoautotróficos e, portanto, são capazes de ter vida livre.
A alternativa está incorreta, pois apenas alguns protozoários podem realizar fotossíntese, e existem várias espécies que são parasitas, as quais obtêm seus nutrientes a partir do tecido do hospedeiro. Além disso, as algas correspondem a organismos fototróficos.
Os vírus e alguns protozoários são parasitas obrigatórios; eles precisam da maquinaria metabólica celular para expressar seus genes.
A alternativa está incorreta, pois, embora a primeira parte da afirmação esteja correta (os vírus e alguns protozoários são, realmente, parasitas obrigatórios), apenas os vírus utilizam maquinaria celular para produzirem proteínas e se multiplicarem. Os protozoários parasitas utilizam o hospedeiro para nutrição.
A reprodução sexuada ocorre apenas no grupo das algas, por meio da produção de gametas; os protozoários, por sua vez, multiplicam-se de forma assexuada (esquizogonia).
A alternativa está incorreta, pois tanto as algas quanto os protozoários se reproduzem de forma sexuada ou assexuada: as algas podem se multiplicar por fragmentação, mas também podem produzir gametas. Os protozoários podem se multiplicar tanto por esquizogonia como por conjugação e pela produção de gametas.
Nome do livro: Microbiologia de Brock
Editora: Artmed
Autor: Michael T. Mandigan, John M. Martinko, Paul V. Dunlap e David P. Clark
ISBN: 978-85-8271-354-9
Microbiologia de Brock é um dos principais livros de Microbiologia, visto que é uma obra completa, com linguagem acessível, conteúdo, esquemas e materiais complementares, voltados para a Microbiologia ambiental.

Nome do livro: Microbiologia
Editora: Artmed
Autor: Gerard J. Tortora, Berdell R. Funke, Christine L. Case
ISBN: 978-85-8271-354-9
Esse é um dos principais livros de Microbiologia, pois, nele, os conteúdos são apresentados de maneira bastante completa e didática, com ilustrações, fotografias e comentários muito esclarecedores. O livro também contém exemplos de aplicações práticas da Microbiologia, além de apresentar problemas que podem ser resolvidos pelos(as) estudantes após o aprendizado de cada capítulo.